ņä£ ļĪĀ
ņŚ¼ņä▒ņŚÉņä£ ĒØöĒĢśĻ▓ī ņ¦äļŗ©ļÉśļŖö ņ£Āļ░®ņĢöņØĆ ļ░£ņĢö Ļ│╝ņĀĢņŚÉņä£ ņłśļ¦ÄņØĆ ņ£ĀņĀäņ×ÉņØś ļ│ĆĒśĢņØ┤ ņ׳ņ¢┤ ņ╣śļŻīĻ░Ć ņēĮņ¦Ć ņĢŖĻ│Ā ņĀäņØ┤ Ļ░ĆļŖźņä▒ņØ┤ ļåÆņØĆ ņ¦łļ│æņ£╝ļĪ£ ļ░®ņé¼ņäĀ ņ╣śļŻī ļśÉļŖö ĒÖöĒĢÖņĀü ņ╣śļŻīļź╝ ļ│æĒ¢ēĒĢ£ ņÖĖĻ│╝ņĀü ņłśņłĀņØ┤ ņĀüņÜ®ļÉśņ¢┤ ņÖöļŗż[1]. ļīĆĒæ£ņĀü ļ░śļĀżļÅÖļ¼╝ņØĖ Ļ░£ņØś Ļ▓ĮņÜ░ņŚÉņä£ļÅä ļ╣łļ▓łĒĢśĻ▓ī ļ░£ļ│æļÉśļŖö ņóģņ¢æ ņżæ ĒĢśļéśņØĖ ņ£ĀņäĀņóģņ¢æņØĆ ļéśņØ┤ ļōĀ ņĢöņ╗Ę Ļ░£ņŚÉņä£ ĒØöĒĢśĻ▓ī ļéśĒāĆļéśļ®░ ĻĘĖ ņżæ ņĀłļ░śņØĆ ņĢģņä▒ņ£╝ļĪ£ ņ¦äļŗ©ļÉśĻ│Ā ņ׳ļŗż[2]. Ļ░£ņØś ņ£ĀņäĀņóģņ¢æņØĆ ņé¼ļ×īĻ│╝ ņ£Āņé¼ĒĢ£ ļ░£ļ│æ ļČĆņ£äļź╝ ļéśĒāĆļé╝ ļ┐Éļ¦ī ņĢäļŗłļØ╝ ņŚŁĒĢÖņĀü, ĒśĢĒā£ĒĢÖņĀü ņĖĪļ®┤ņŚÉņä£ļÅä ņŚ¼ņä▒ņØś ņ£Āļ░®ņĢöĻ│╝ ņ£Āņé¼ņä▒ņØä ņ¦ĆļŗłĻ│Ā ņ׳ņ¢┤ ņ£Āļ░®ņĢö ņŚ░ĻĄ¼ļČäņĢ╝ņØś ņŗżĒŚśļ¬©ļŹĖļĪ£ņä£ļÅä ĒÖ£ņÜ®ļÉśĻ│Ā ņ׳ļŗż[3,4].
Ļ░£ņŚÉņä£ ņĢöņØä ņ╣śļŻīĒĢśļŖö ļ░®ļ▓Ģņ£╝ļĪ£ļŖö ņŻ╝ļĪ£ ņÖĖĻ│╝ņĀü ņłśņłĀņØä ņé¼ņÜ®ĒĢ┤ņÖöļŗż. 1946ļģäņŚÉ ļ░▒Ēśłļ│æ ņ¦äļŗ©ņØä ļ░øņØĆ Ļ░£ļź╝ ļīĆņāüņ£╝ļĪ£ ĒÖöĒĢÖņĀü ņ╣śļŻīĻ░Ć ņŗ£ļÅäļÉśņŚłĻ│Ā[5], ņØ┤Ēøä ņĀÉņ░©ņĀüņ£╝ļĪ£ Ļ░üņóģ ņĢöņŚÉ ĒÖöĒĢÖņĀü ņ╣śļŻīĻ░Ć ņĀüņÜ®ļÉśĻĖ░ ņŗ£ņ×æĒĢśņśĆņ£╝ļéś[6], ĒÖśņČĢņØä ļīĆņāüņ£╝ļĪ£ ņןĻĖ░Ļ░ä ņ╣śļŻīļź╝ ņ¦ĆņåŹĒĢśļŖö Ļ▓ĮņÜ░ļŖö ļō£ļ¼╝Ļ│Ā ņØ┤ļ¤¼ĒĢ£ ĒÖöĒĢÖņĀü ņ╣śļŻīņØś ĒÜ©ļŖźņØä ļÆĘļ░øņ╣©ĒĢĀ ļ¦īĒĢ£ ņłśņØśĒĢÖņĀü ņ×ÉļŻī ļČĆņĪ▒ņØś ĒĢ£Ļ│äĻ░Ć ņ׳ņŚłļŗż[7]. ņØ┤Ēøä ņ▓┤ļé┤ ļ®┤ņŚŁņ▓┤Ļ│äĻ░Ć ņĢöņØś ņä▒ņןņØä ņĪ░ņĀłĒĢśĻ│Ā ņÖäĒÖöĒĢĀ ņłś ņ׳ļŗżļŖö ņŚ░ĻĄ¼ Ļ▓░Ļ│╝ļōżņØ┤ ļ│┤Ļ│ĀļÉśļ®┤ņä£ ņĄ£ĻĘ╝ ļīĆĒæ£ņĀüņØĖ ļ®┤ņŚŁņäĖĒżņØĖ ņ×ÉņŚ░ņé┤ĒĢ┤ņäĖĒż(natural killer cells, NKņäĖĒż)ļéś TņäĖĒż ļō▒ņØä Ēæ£ņĀüņ£╝ļĪ£ ĒĢśļŖö ļ®┤ņŚŁ ņÜöļ▓ĢņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼ļōżņØ┤ ĒÖ£ļ░£ĒĢśĻ▓ī ņ¦äĒ¢ēļÉśĻ│Ā ņ׳ņ£╝ļ®░, ņłśņØśļČäņĢ╝ņŚÉņä£ļÅä ļÅÖļ¼╝ņØś ļŗżņ¢æĒĢ£ ņ¦łļ│æņØä ļīĆņāüņ£╝ļĪ£ ļ®┤ņŚŁ ņÜöļ▓ĢņŚÉ ļīĆĒĢ£ ĻĖ░ņ┤ł ļ░Å ņ×äņāü Ļ┤ĆļĀ© ņĀüņÜ® ņŚ░ĻĄ¼ļōżņØ┤ ņ¦äĒ¢ēļÉśĻ│Ā ņ׳ļŗż[4].
Ļ░üņóģ ļ®┤ņŚŁņäĖĒż ņżæ NKņäĖĒżļŖö ņāØņ▓┤ ļé┤ ņĢö ļ░£ņāØņØä ņ¢ĄņĀ£ĒĢśļŖö ņĄ£ņĀäņäĀņØś ĒĢĄņŗ¼ ļ®┤ņŚŁņäĖĒżļĪ£ņŹ© ĒĢŁņøÉĒŖ╣ņØ┤ņĀüņØĖ ņłśņÜ®ņ▓┤ļź╝ Ļ░Ćņ¦ä TņäĖĒż, BņäĖĒżņÖĆ ļŗ¼ļ”¼ ļŗżņ¢æĒĢ£ ņäĀņ▓£ļ®┤ņŚŁ ņłśņÜ®ņ▓┤ļź╝ ņäĖĒż Ēæ£ļ®┤ņŚÉ ļ░£ĒśäĒĢśĻ│Ā ņØ┤ļōżņØä ĒåĄĒĢ┤ ņĀĢņāüņäĖĒżņÖĆ ņĢöņäĖĒżļź╝ ĻĄ¼ļČäĒĢśļ®░, ĒŹ╝Ēżļ”░(perforin) ļ░Å ĻĘĖļ×£ņ×Éņ×ä(granzyme B)ņØä ļČäļ╣äĒĢśņŚ¼ ņ”ēĻ░üņĀüņ£╝ļĪ£ ņĢöņäĖĒżļź╝ ņé¼ļ®ĖĒĢśļŖö ĒŖ╣ņ¦ĢņØä Ļ░Ćņ¦äļŗż. ņØ┤ļ¤¼ĒĢ£ NKņäĖĒżļŖö ņĢöņØś ļ░£ņāØ, ņĀäņØ┤, ņ×¼ļ░£ņŚÉ ņżæņÜöĒĢ£ ņŚŁĒĢĀņØä ĒĢśļŖö ņĢöņżäĻĖ░ņäĖĒż(cancer stem cells)ļź╝ ĒÜ©Ļ│╝ņĀüņ£╝ļĪ£ ņĀ£Ļ▒░ĒĢĀ ņłś ņ׳ņ¢┤ ĒĢŁņĢö ļ®┤ņŚŁņÜöļ▓ĢņŚÉņä£ ļ¦ÄņØĆ ņןņĀÉņØä Ļ░¢Ļ│Ā ņ׳ļŗż. ļŗżņ¢æĒĢ£ ņĢö ĒÖśņ×ÉņŚÉņä£ NKņäĖĒżņØś ĻĖ░ļŖź Ļ▓░ĒĢ©ņØ┤ ļ│┤Ļ│ĀļÉśĻ│Ā ņ׳ņ£╝ļ®░, ņØ┤ ĻĖ░ļŖź Ļ▓░ĒĢ©ņØś ņĀĢļÅäĻ░Ć Ē¢źĒøä ņ×äņāüņŚÉņä£ņØś ņśłĒøäņÖĆ ļ░ĆņĀæĒĢ£ ņāüĻ┤ĆĻ┤ĆĻ│äĻ░Ć ņ׳ļŗżļŖö Ļ▓āņØ┤ ļ│┤Ļ│ĀļÉśņŚłļŗż. Ēśäņ×¼ NKņäĖĒżļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņé¼ļ×īņØś ņĢöņØä ņ╣śļŻīĒĢśļĀżļŖö ļŗżņ¢æĒĢ£ ņŗ£ļÅäļōżņØ┤ ņ׳ņ£╝ļ®░[8], Ļ░£ņŚÉņä£ļÅä ņŚ░ĻĄ¼Ļ░Ć ņ¦äĒ¢ē ņżæņØ┤ļŗż[9].
NKņäĖĒżņØś ļīĆĒæ£ņĀü ļ¦łņ╗żņØĖ NKp46 (NCR1)ņŚÉ ĻĖ░ņ┤łĒĢśņŚ¼ ļ│╝ ļĢī, Ļ░£ NKņäĖĒżļŖö ņĀäņ▓┤ ļ¦Éņ┤łĒśłņĢĪ ļŗ©ĒĢĄņäĖĒżņŚÉņä£ ņĢĮ 5% ņĀĢļÅäļĪ£ ņĪ┤ņ×¼ĒĢśņŚ¼ ņé¼ļ×ī NKņäĖĒż ļ╣äņ£©(10~15%)ņŚÉ ļ╣äĒĢ┤ ĻĘĖ ļ╣äņ£©ņØ┤ ĒśäņĀĆĒ׳ ņĀüļŗż[7]. ņé¼ļ×īņŚÉņä£ CD5 ĒĢŁņøÉņØĆ ņĀĢņāü TņäĖĒżņŚÉ ņĪ┤ņ×¼ĒĢśļ®░ NKņäĖĒżņŚÉņä£ļŖö ļ░£ĒśäĒĢśņ¦Ć ņĢŖļŖöļŗżĻ│Ā ļ│┤Ļ│ĀļÉśņŚłņ£╝ļ®░[10], Huang ļō▒ņØĆ CD5 ņØīņä▒ ņäĖĒżĻ░Ć Ļ░£ NKņäĖĒżņØś ĒŖ╣ņä▒Ļ│╝ ļ░ĆņĀæĒĢśĻ▓ī Ļ┤ĆļĀ©ļÉśņ¢┤ ņ׳ņØīņØä ļ│┤Ļ│ĀĒĢśņśĆļŗż[11]. Canter ļō▒ļÅä Ļ│©ņ£ĪņóģņØä Ļ░Ćņ¦ä Ļ░£ņŚÉĻ▓ī ļ░®ņé¼ņäĀ ņ╣śļŻī Ēøä CD5dim, NKp46+ ņäĖĒżļź╝ Ļ░£ņŚÉĻ▓ī Ēł¼ņŚ¼ĒĢśņŚ¼ NKņäĖĒżņØś ĒÖ£ņä▒ĒÖö ņ”ØĻ░Ć ļ░Å ņóģņ¢æ ņä▒ņן ņ¦ĆņŚ░ņØä ļ│┤Ļ│ĀĒĢ£ ļ░ö ņ׳ļŗż[9].
ņé¼ļ×īņŚÉņä£ ņ£Āļ░®ņĢöņØä ņ¦äļŗ©ĒĢśļŖö ļīĆĒæ£ņĀü ļ░öņØ┤ņśżļ¦łņ╗żļĪ£ļŖö estrogen receptor (ER), progesterone receptor (PR), human epidermal growth factor receptor type-2 (HER-2) ļō▒ņØ┤ ņ׳ņ£╝ļ®░, ņ£Āļ░®ņĢöņØś ņĢĮ 70% ņĀĢļÅäļŖö ņĢöņäĖĒż ņä▒ņן ņ┤ēņ¦äņŚÉ ņØ┤ļ¤¼ĒĢ£ ĒśĖļź┤ļ¬¼ļōżņØ┤ ņśüĒ¢źņØä ļ»Ėņ╣£ļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ļŗż[1]. Ļ░£ņØś ņ£ĀņäĀņóģņ¢æņŚÉņä£ļŖö ņØ┤ļ¤¼ĒĢ£ ĒśĖļź┤ļ¬¼ ņłśņÜ®ņ▓┤ļ┐Éļ¦ī ņĢäļŗłļØ╝ proliferative cell nuclear antigen (PCNA), tumor suppressor protein p53 (p53), E-cadherin, endothelial growth factor ļ░Å ņĢöņżäĻĖ░ņäĖĒż ĒŖ╣ņØ┤ļ¦łņ╗ż ļō▒ņØ┤ ņĢīļĀżņĀĖ ņ׳ņ£╝ļ®░, ņØ┤ļ¤¼ĒĢ£ ļ¦łņ╗żļōżņØĆ ņóģņ¢æņØś ņĢģņä▒ļÅäņŚÉ ļö░ļØ╝ ļŗżļźĖ ļ░£ĒśäņØä ļéśĒāĆļé┤ļ»ĆļĪ£ ĒĢŁņĢö ļ®┤ņŚŁņÜöļ▓ĢņŚÉ ļīĆĒĢ£ ļ░śņØæņØĆ ņ£ĀņäĀņóģņ¢æĻ│╝ Ļ┤ĆļĀ©ļÉ£ ĒŖ╣ņØ┤ļ¦łņ╗żņŚÉ ļö░ļØ╝ ļŗżļź┤Ļ▓ī Ļ▓░ņĀĢļÉĀ ņłś ņ׳ļŗż[12]. ņØ┤ļ¤¼ĒĢ£ ļ░öņØ┤ņśżļ¦łņ╗żļŖö ĒŖ╣ņĀĢ ņ╣śļŻīņŚÉ ļīĆĒĢ£ ņ×Āņ×¼ņĀüņØĖ ļ░śņØæņØä Ļ▓░ņĀĢĒĢśļŖö ļŹ░ņŚÉ ņ£ĀņÜ®ĒĢ£ ņĀĢļ│┤ļź╝ ņĀ£Ļ│ĄĒĢśĻ│Ā ņä▒Ļ│ĄņĀüņØĖ ņ╣śļŻī ņĀäļץņØä ņ£äĒĢ┤ ņżæņÜöĒĢśĻ▓ī ņé¼ņÜ®ļÉ£ļŗż[13].
ļ│Ė ņŚ░ĻĄ¼ļŖö ņä£ļĪ£ ļŗżļźĖ ļ░öņØ┤ņśżļ¦łņ╗żļź╝ ļ░£ĒśäĒĢśļŖö Ļ░£ ņ£ĀņäĀņóģņ¢æ ņ£Āļל 2Ļ░Ćņ¦Ć ņäĖĒżļź╝ ļīĆņāüņ£╝ļĪ£ Ļ░£ ļ¦Éņ┤łĒśłņĢĪņ£╝ļĪ£ļČĆĒä░ ļČäļ”¼ĒĢ£ CD5 ņØīņä▒ ņäĖĒżļź╝ ņ▓┤ņÖĖņŚÉņä£ ņāüļŗ╣ļ¤ē ņ”ØĒÅŁ ļ░Å NKņäĖĒżļĪ£ ĒÖ£ņä▒ĒÖöĒĢ£ ļŗżņØī, Ļ░£ ņ£ĀņäĀņóģņ¢æņäĖĒż(Ēæ£ņĀüņäĖĒż)ņÖĆ Ļ│Ąļ░░ņ¢æĒĢśņŚ¼ Ēæ£ņĀüņäĖĒżņŚÉ ļīĆĒĢ£ NKņäĖĒżņØś ļÅģņä▒ ļ░Å ĻĘĖ ņ░©ņØ┤ļź╝ ļČäņäØĒĢśņśĆņ£╝ļ®░, Ē¢źĒøä Ļ░£ ņ£ĀņäĀņóģņ¢æ ļ®┤ņŚŁņäĖĒż ņ╣śļŻī ņŚ░ĻĄ¼ņŚÉ ļÅäņøĆņØ┤ ļÉśĻ│Āņ×É ĒĢśņśĆļŗż.
ņ×¼ļŻī ļ░Å ļ░®ļ▓Ģ
Ļ░£ ņ£ĀņäĀņóģņ¢æņäĖĒż ļ░░ņ¢æ
Ļ░£ ņ£ĀņäĀņóģņ¢æņäĖĒżņØĖ REM134 (ECACC, UK)ņÖĆ CF41.Mg (ATCC, USA)ļź╝ ņé¼ņÜ®ĒĢśņśĆļŗż. REM134 ņäĖĒżņÖĆ CF41.Mg ņäĖĒżļŖö 10% fetal bovine serum (FBS; Gibco, USA), 100 U/mL penicillin, 100 ╬╝g/mL streptomycin ļ░Å 1├Ś glutamax (Gibco)Ļ░Ć ņ▓©Ļ░ĆļÉ£ DulbeccoŌĆÖs modified EagleŌĆÖs medium (Gibco)ņØä ņé¼ņÜ®ĒĢśņŚ¼ 37┬░C, 5% CO2 ņĪ░Ļ▒┤ņØś ņäĖĒż ļ░░ņ¢æĻĖ░ņŚÉņä£ ļ░░ņ¢æĒĢśņśĆļŗż.
Ļ░£ NKņäĖĒż ļČäļ”¼ ļ░Å ņ”ØņŗØ
Ļ▒┤Ļ░ĢĒĢ£ ņĢöņ╗Ę Ļ░£(female beagle)ņØś ļ¦Éņ┤łĒśłņĢĪ(Genia Inc., Korea) ļŗ©ĒĢĄņäĖĒż(peripheral blood mononuclear cells, PBMCs)ļĪ£ļČĆĒä░ NKņäĖĒżļōżļ¦ī ņäĀĒāØņĀüņ£╝ļĪ£ ļ░░ņ¢æĒĢśĻ│Āņ×É PBMCsņŚÉņä£ CD5ņØīņä▒ ņäĖĒżļ¦īņØä ļČäļ”¼ĒĢśņśĆļŗż. ļ¦Éņ┤łĒśłņĢĪņØä PBSņÖĆ 1:1ļĪ£ ĒؼņäØĒĢśņśĆņ£╝ļ®░, Ficoll-Paque (GE Healthcare, Sweden)ņØä ņé¼ņÜ®ĒĢśņŚ¼ ĒؼņäØļÉ£ ļ¦Éņ┤łĒśłņĢĪņ£╝ļĪ£ļČĆĒä░ PBMCsļź╝ ļČäļ”¼ĒĢśņŚ¼ 2ĒÜī ņäĖņ▓ÖĒĢśņśĆļŗż. PBMCsļź╝ ļīĆņāüņ£╝ļĪ£ CD5 (MCA1037PE, BioRad, USA) ĒĢŁņ▓┤ļź╝ 30ļČäĻ░ä ņŚ╝ņāēĒĢ£ ļŗżņØī, anti-PE microbeads (Miltenyi Biotec, USA)ļĪ£ 2ņ░© ņŚ╝ņāēĒĢśņŚ¼ MACS ņŗ£ņŖżĒģ£(Miltenyi Biotec)ņ£╝ļĪ£ CD5 ĒĢŁņ▓┤Ļ░Ć ļČĆņ░®ļÉśņ¦Ć ņĢŖņØĆ ņäĖĒżļź╝ ņłśņ¦æĒĢśņśĆļŗż. ņłśņ¦æĒĢ£ ņäĖĒżļź╝ ņ£ĀņäĖĒż ļČäņäØ(FACS Cabilur, USA)ņØä ĒåĄĒĢ┤ CD5 ņØīņä▒ ņäĖĒżņ×äņØä ĒÖĢņØĖĒĢśņśĆļŗż. ļśÉĒĢ£, NKņäĖĒżļ¦īņØä ņäĀĒāØņĀüņ£╝ļĪ£ ņ”ØņŗØĒĢśĻĖ░ ņ£äĒĢ┤ ņé¼ļ×ī ĒśłņĢĪņĢö ņäĖĒżņŻ╝ņØĖ K562ļź╝ ņśüņ¢æņäĖĒż(feeder cells)ļĪ£ ņé¼ņÜ®ĒĢśņśĆļŗż. K562ņŚÉ ļ░®ņé¼ņäĀ Ļ░Éļ¦łņäĀ(100 Gy)ņØä ņĪ░ņé¼ĒĢśņŚ¼ ņäĖĒż ļČäņŚ┤ņØä ņĀĆĒĢ┤ĒĢ£ ļŗżņØī 6 well plateņŚÉņä£ K562 (5 ├Ś 105Ļ░£/well)ņÖĆ CD5 ņØīņä▒ ņäĖĒż(1 ├Ś 106Ļ░£/well)ļź╝ 10% FBS (Gibco), 100 U/mL penicillin, 100 ╬╝g/mL streptomycin, 500 U/mL human IL-2 (R&D, USA), 10 ng/mL human IL-15 (R&D) ļ░Å 5 ng/mL canine IL-21 (R&D)Ļ░Ć ņ▓©Ļ░ĆļÉ£ RPMI, GlutaMAXTM (Gibco) ļ░░ņ¢æņĢĪņŚÉņä£ 7ņØ╝ Ļ░äĻ▓®ņ£╝ļĪ£ Ļ│äļīĆĒĢśņŚ¼ ņ┤Ø 21ņØ╝Ļ░ä 37┬░C, 5% CO2 ņĪ░Ļ▒┤ņØś ņäĖĒż ļ░░ņ¢æĻĖ░ņŚÉņä£ ļ░░ņ¢æĒĢśņśĆļŗż.
ņĀĢļ¤ēņĀü ņżæĒĢ®ĒÜ©ņåīņŚ░ņćäļ░śņØæ(quantitative real-time reverse transcriptase-polymerase chain reaction, qPCR)ņØä ņØ┤ņÜ®ĒĢ£ ņ£ĀņĀäņ×É ļČäņäØ
ņäĖĒżņŚÉņä£ RNeasy mini kit (Qiagen, USA)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ RNAļź╝ ņČöņČ£ĒĢśĻ│Ā Nano-drop 1000 (Thermo, USA)ņØä ņé¼ņÜ®ĒĢśņŚ¼ RNA ņāüĒā£ņÖĆ ļåŹļÅäļź╝ ņĖĪņĀĢĒĢ£ ļŗżņØī, GoScriptTM Reverse Transcriptase (Promega, USA)ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ cDNA ĒĢ®ņä▒ĒĢśņśĆļŗż. qPCRņØĆ 96 well plateņŚÉ LightCycler┬« 480 SYBR Green I Master Mix (Roche Diagnostics, USA)ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ pre-denaturation (95┬░C, 10ļČä), annealing (60┬░C, 10ņ┤ł), elongation (72┬░C, 10ņ┤ł)ņØä 45 cycle ļ░śļ│ĄĒĢ£ Ēøä 2ŌłÆ╬öCt Ļ│äņé░ļ▓ĢņØä ņØ┤ņÜ®ĒĢśņŚ¼ melting curveļź╝ ļČäņäØĒĢśņśĆņ£╝ļ®░, ╬▓-actinņØä reference geneņ£╝ļĪ£ ņé¼ņÜ®ĒĢśņŚ¼ qPCR Ļ▓░Ļ│╝ļź╝ ļČäņäØĒĢśņśĆļŗż. ņé¼ņÜ®ļÉ£ primerņØś ņĀĢļ│┤ņÖĆ ņĪ░Ļ▒┤ņØĆ Table 1ņŚÉ ļéśĒāĆļé┤ņŚłļŗż.
Enzyme-linked immunosorbent assay (ELISA)ļź╝ ĒåĄĒĢ£ IFN-╬│ ļČäņäØ
Ēæ£ņĀüņäĖĒżņØĖ REM134ņÖĆ CF41.MgņŚÉ ļīĆĒĢ£ Ļ░£ NKņäĖĒżņØś ĒÖ£ņä▒ņØä ļČäņäØĒĢśĻ│Āņ×É IFN-╬│ ELISA kit (DY781B, R&D systems, USA)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ĒÖ£ņä▒ĒÖöļÉ£ NKņäĖĒżņØś ņŻ╝ņÜöĒĢ£ ĒŖ╣ņ¦ĢņØĖ IFN-╬│ņØś ņāØņä▒ņØä ļČäņäØĒĢśņśĆļŗż. REM134, CF41.MgņÖĆ NK ņäĖĒżļź╝ 1:10 (2 ├Ś 105 : 2 ├Ś 106) ļ╣äņ£©ļĪ£ RPMI, GlutaMAXTM (Gibco)ņŚÉ 10% FBS (Gibco), 100 U/mL penicillin ļ░Å 100 ╬╝g/mL streptomycinņØ┤ ņ▓©Ļ░ĆļÉ£ ņäĖĒż ļ░░ņ¢æņĢĪņ£╝ļĪ£ 37┬░C, 5% CO2 ņĪ░Ļ▒┤ņØś ņäĖĒż ļ░░ņ¢æĻĖ░ņŚÉņä£ 24ņŗ£Ļ░ä Ļ│Ąļ░░ņ¢æĒĢ£ ļŗżņØī, ņāüņĖĄņĢĪņØä ņłśņ¦æĒĢśņŚ¼ IFN-╬│ņØś ļåŹļÅäļź╝ ņĖĪņĀĢĒĢśņśĆļŗż.
Ēæ£ņĀüņäĖĒżņŚÉ ļīĆĒĢ£ NKņäĖĒż ļÅģņä▒ ļČäņäØ
Ēæ£ņĀüņäĖĒżņØĖ REM134ņÖĆ CF41.MgņŚÉ ļīĆĒĢ£ Ļ░£ NKņäĖĒżņØś ļÅģņä▒ņØä ļČäņäØĒĢśĻ│Āņ×É lactase dehydrogenase (LDH) ļ░®ņČ£ņØä ņØ┤ņÜ®ĒĢ£ non-radioactive cytotoxicity assay kit (Promega)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ NKņäĖĒżņØś Ēæ£ņĀüņäĖĒżņŚÉ ļīĆĒĢ£ ņäĖĒż ļÅģņä▒ņØä ļČäņäØĒĢśņśĆļŗż. NKņäĖĒżņÖĆ Ēæ£ņĀüņäĖĒżļź╝ 25:1, 12.5:1, 6.25:1, 3.13:1, 1.56:1 ļ╣äņ£©ļĪ£ U bottom 96 well plateņŚÉ ņäĖĒżļź╝ ļČäņŻ╝ĒĢśņŚ¼ NKņäĖ ĒżņÖĆ Ēæ£ņĀüņäĖĒżļź╝ 37┬░C, 5% CO2 ņĪ░Ļ▒┤ņØś ņäĖĒż ļ░░ņ¢æĻĖ░ņŚÉņä£ 4ņŗ£Ļ░ä Ļ│Ąļ░░ņ¢æĒĢ£ ļŗżņØī ĒØĪĻ┤æļÅä 490 nmņŚÉņä£ LDHļź╝ ņĖĪņĀĢĒĢśņśĆļŗż.
Ļ▓░ Ļ│╝
Ļ░£ ņ£ĀņäĀņóģņ¢æņäĖĒż ļ¬©ņ¢æ ļ░Å ĒŖ╣ņØ┤ļ¦łņ╗ż ļ░£Ēśä
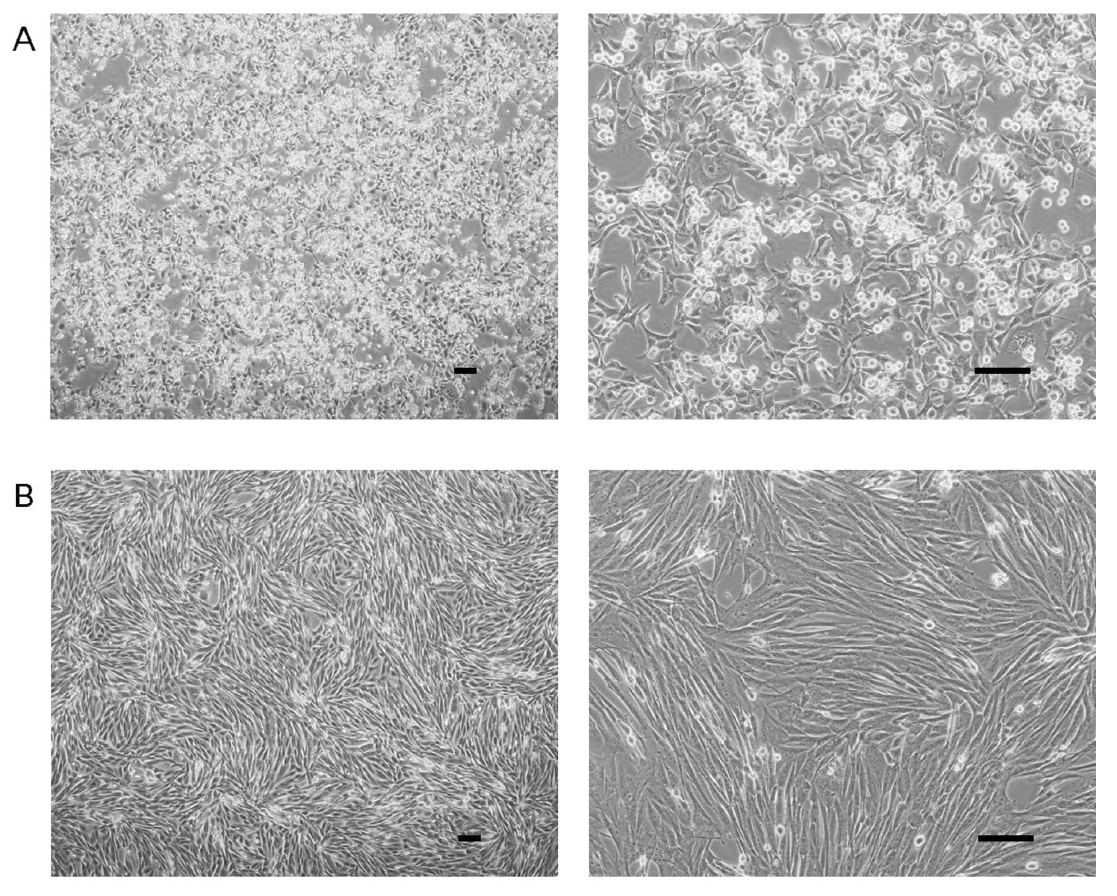
Ļ░£ ņ£ĀņäĀņóģņ¢æņäĖĒżņØĖ REM134ņÖĆ CF41.Mg 1 ├Ś 106Ļ░£ļź╝ Ļ░üĻ░ü T75 ĒöīļØ╝ņŖżĒü¼ņŚÉ ļČĆņ░®ĒĢśņśĆļŗż. ņĄ£ņ┤ł ļČĆņ░® ņØ┤Ēøä REM134ļŖö ņāüĒö╝ņäĖĒż ĒśĢĒā£ļĪ£ ņ×ÉļØ╝ļŖö Ļ▓āņØä ĒÖĢņØĖĒĢśņśĆĻ│Ā CF41.MgļŖö ņżæĻ░ä ņŚĮņäĖĒżņÖĆ ņ£Āņé¼ĒĢ£ ĒśĢĒā£ļĪ£ ņ×ÉļØ╝ļŖö Ļ▓āņØä ĒÖĢņØĖĒĢśņśĆņ£╝ļ®░, 2Ļ░Ćņ¦Ć ņäĖĒż ļ¬©ļæÉ 3ņØ╝ Ēøä ņĢĮ 80% confluencyņŚÉ ļÅäļŗ¼ĒĢśņśĆļŗż(Fig. 1).
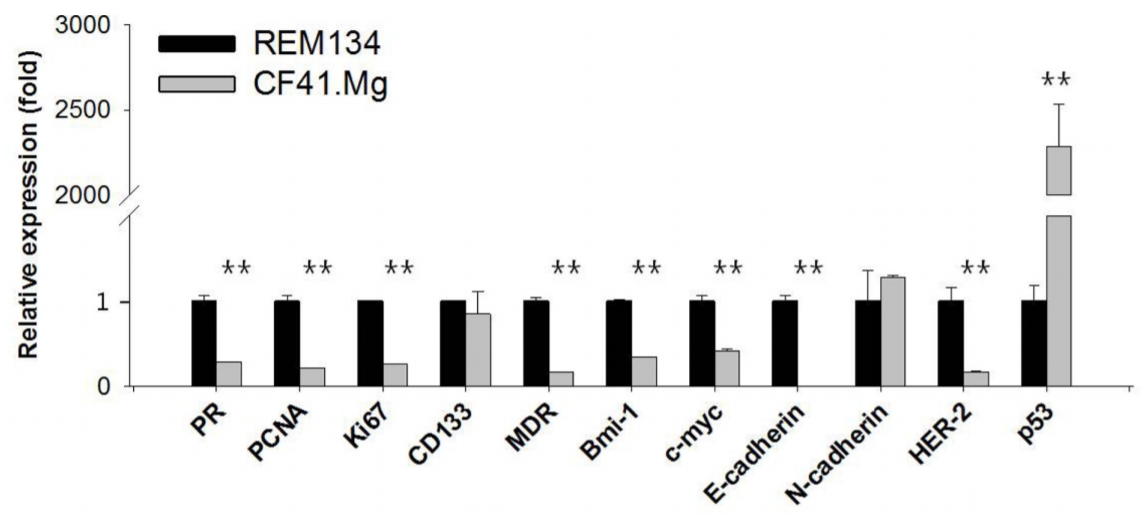
REM134ņÖĆ CF41.MgņŚÉņä£ ņ£ĀņäĀņóģņ¢æ ĒŖ╣ņØ┤ ļ░öņØ┤ņśżļ¦łņ╗żļōżņØś ļ░£ĒśäņØä ĒÖĢņØĖĒĢśĻ│Āņ×É PR, CD133, MDR (multiple drug resistance), Bmi-1, c-myc, E-cadherin, N-cadherin, HER-2, p53, PCNA ļ░Å Ki67ņØś ļ░£ĒśäņØä ļČäņäØĒĢśņśĆļŗż. ĻĘĖ Ļ▓░Ļ│╝, CF41.MgņØĆ REM134ņŚÉ ļ╣äĒĢ┤ ņ£ĀņØśņĀüņ£╝ļĪ£ Bmi-1, MDR, c-myc, HER-2, PCNA ļ░Å Ki67ņØś ļ░£ĒśäņØ┤ ļé«ņĢśĻ│Ā CD133, N-cadherinņØś ļ░£ĒśäņŚÉļŖö Ēü░ ņ░©ņØ┤Ļ░Ć ņŚåņŚłņ£╝ļ®░, p53Ļ│╝ E-cadherinņØś ļ░£ĒśäņØĆ ņśżĒ׳ļĀż ņ£ĀņØśņĀüņ£╝ļĪ£ ļåÆņØīņØä ĒÖĢņØĖĒĢśņśĆļŗż(Fig. 2).
NKņäĖĒż ņ▓┤ņÖĖ ņ”ØņŗØ ļ░Å ĒÖ£ņä▒ĒÖö
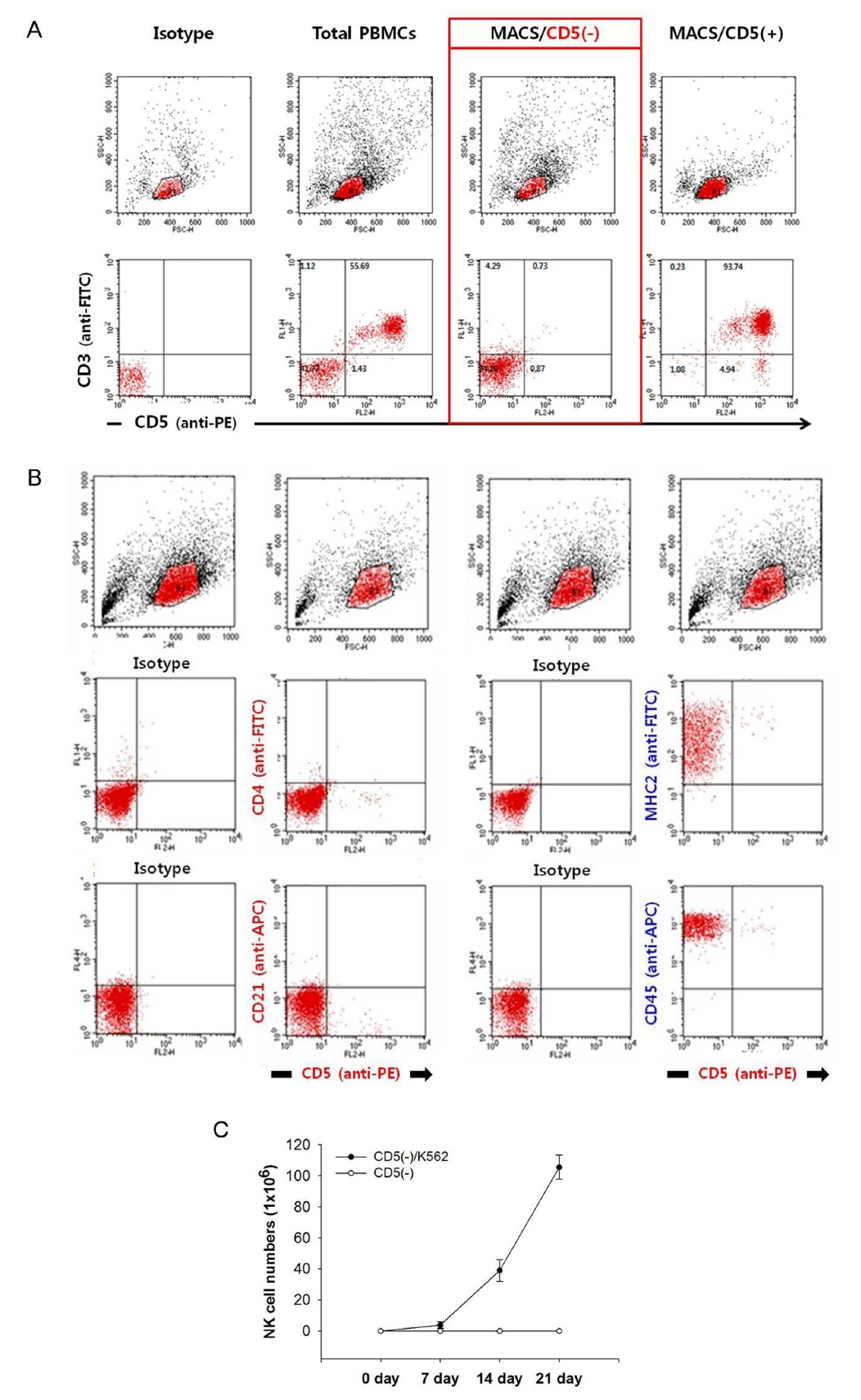
Ļ░£ PBMCsļĪ£ļČĆĒä░ MACS systemņ£╝ļĪ£ CD5 ņØīņä▒ ņäĖĒżļź╝ ļČäļ”¼ĒĢśņŚ¼ ņ£ĀņäĖĒż ļČäņäØņØä ĒåĄĒĢ┤ ĻĘĖ Ļ▓░Ļ│╝ļź╝ ĒÖĢņØĖĒĢśņśĆļŗż(Fig. 3A and B). ļŗżņØīņ£╝ļĪ£ Ļ░£ CD5 ņØīņä▒ ņäĖĒżņÖĆ Ļ░Éļ¦łņäĀ(100 Gy)ņØ┤ ņĪ░ņé¼ļÉ£ K562 ņäĖĒżņŚÉ IL-2, IL-15 ļ░Å IL-21ņØä ņ▓©Ļ░ĆĒĢśņŚ¼ 21ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņŚ¼ NKņäĖĒż ņ▓┤ņÖĖ ņ”ØņŗØ ļ░Å ĒÖ£ņä▒ĒÖöļÉ£ NKņäĖĒżņØś ņłśņÜ®ņ▓┤ ļ░£ĒśäņØä ļČäņäØĒĢśņśĆļŗż. ĻĘĖ Ļ▓░Ļ│╝, ņĄ£ņ┤łņØś CD5 ņØīņä▒ ņäĖĒżņŚÉ ļ╣äĒĢ┤ 21ņØ╝Ļ░ä ļ░░ņ¢æņØä Ļ▒░ņ╣£ CD5 ņØīņä▒ ņäĖĒżļŖö ņĢĮ 100ļ░░ ņØ┤ņāü ņäĖĒż ņłśĻ░Ć ņ”ØņŗØĒĢśņśĆņ£╝ļ®░(Fig. 3C), ĒÖ£ņä▒ĒÖöļÉ£ NKņäĖĒżņØś ĒŖ╣ņØ┤ļ¦łņ╗żņØĖ NKp30 (17.6 ┬▒ 1.4), NKp44 (24.7 ┬▒ 2.0), NKp46 (522.9 ┬▒ 22.8), NKG2D (63.6 ┬▒ 2.2), CD244 (1.3 ┬▒ 0.0), perforin (118.6 ┬▒ 10.6), granzyme B (229.2 ┬▒ 14.6) ļ░Å TNF-╬▒ (6.0 ┬▒ 0.6)ņØś ļ░£ĒśäņØ┤ ĒśäņĀĆĒĢśĻ▓ī ņ”ØĻ░ĆĒĢśņśĆņØīņØä ĒÖĢņØĖĒĢśņśĆļŗż(Fig. 4).
ĒÖ£ņä▒ĒÖöļÉ£ NKņäĖĒżņØś IFN-╬│ ņāØņä▒
IFN-╬│ ņāØņä▒ ņ”ØĻ░ĆļŖö ĒÖ£ņä▒ĒÖöļÉ£ NKņäĖĒżĻ░Ć Ēæ£ņĀüņäĖĒżļź╝ ņØĖņ¦ĆĒĢśņŚ¼ ļéśĒāĆļéśļŖö ņŻ╝ņÜöĒĢ£ ĒŖ╣ņ¦Ģ ņżæ ĒĢśļéśņØ┤ļŗż. CD5 ņØīņä▒ ņäĖĒżļź╝ 21ņØ╝Ļ░ä NKņäĖĒżļĪ£ ĒÖ£ņä▒ĒÖöĒĢśņŚ¼ ĒŖ╣ņØ┤ļ¦łņ╗żņØś ĒśäņĀĆĒĢ£ ļ░£ĒśäņØä ĒÖĢņØĖĒĢśņśĆņ£╝ļ»ĆļĪ£ ĒÖ£ņä▒ĒÖöļÉ£ NKņäĖĒżņÖĆ Ēæ£ņĀüņäĖĒżļź╝ 24ņŗ£Ļ░ä Ļ│Ąļ░░ņ¢æĒĢśņŚ¼ IFN-╬│ ņāØņä▒ņØ┤ ņ”ØĻ░ĆĒĢśļŖöņ¦Ćļź╝ ļČäņäØĒĢśņśĆļŗż. ĻĘĖ Ļ▓░Ļ│╝, ĒÖ£ņä▒ĒÖöļÉ£ NKņäĖĒżņÖĆ Ēæ£ņĀüņäĖĒżņØĖ REM134 (181.1 ┬▒ 10.0 pg/mL), CF41.Mg (172.6 ┬▒ 1.2 pg/mL)ņÖĆ ĒĢ©Ļ╗ś Ļ│Ąļ░░ņ¢æĒĢ£ ņāüņĖĄņĢĪņŚÉņä£ NKņäĖĒż ļŗ©ļÅģ ļ░░ņ¢æĻĄ░(78.9 ┬▒ 0.7 pg/mL) ļ│┤ļŗż 2ļ░░ ņØ┤ņāü ļåÆņØĆ IFN-╬│ ņāØņä▒ņØä ĒÖĢņØĖĒĢśņśĆļŗż(Fig. 5).
NKņäĖĒżņØś Ēæ£ņĀüņäĖĒżņŚÉ ļīĆĒĢ£ ņäĖĒż ļÅģņä▒
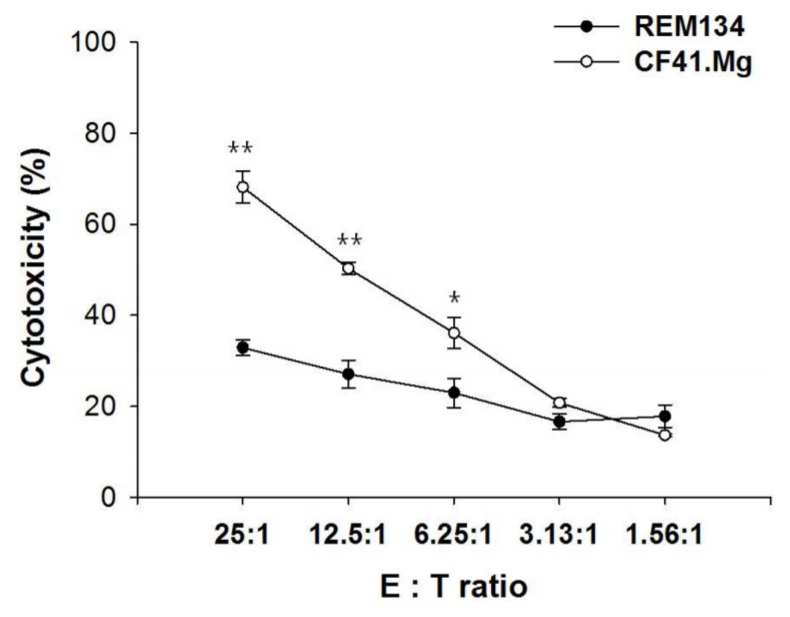
NKņäĖĒżņØś Ēæ£ņĀüņäĖĒż(target cells, T) REM134, CF41.MgņŚÉ ļīĆĒĢ£ ņäĖĒż ļÅģņä▒ņØä ļČäņäØĒĢśĻ│Āņ×É ĒÖ£ņä▒ĒÖöļÉ£ NKņäĖĒżļź╝ ņ×æļÅÖ ņäĖĒż(effector cells, E)ļĪ£ ĒÖ£ņÜ®ĒĢśņŚ¼ E:T ļ╣äņ£©ņØ┤ 25:1-1.56:1ņØ┤ ļÉśļÅäļĪØ Ēś╝ĒĢ®ĒĢśņŚ¼ ņäĖĒż ļÅģņä▒ņØä ļČäņäØĒĢśņśĆļŗż. ĻĘĖ Ļ▓░Ļ│╝ NKņäĖĒżļŖö Ēæ£ņĀüņäĖĒżņØĖ REM134, CF41.MgņŚÉņä£ ņäĖĒż ļÅģņä▒ņØä ļéśĒāĆļé┤ņŚłļŖöļŹ░, E:T ļ╣äņ£©ņØ┤ 25:1ņØĖ Ļ▓ĮņÜ░, REM134ļŖö 32.9%, CF41.MgņØĆ 68.2%ņØś ņäĖĒż ļÅģņä▒ņØ┤ ĒÖĢņØĖļÉśņŚłļŗż. ņ”ē, NKņäĖĒżņØś 2Ļ░Ćņ¦Ć Ēæ£ņĀüņäĖĒżņŚÉ ļīĆĒĢ£ ņäĖĒż ļÅģņä▒ņØä ļ╣äĻĄÉĒĢ£ Ļ▓░Ļ│╝, REM134ņŚÉ ļ╣äĒĢ┤ CF41.MgņŚÉņä£ ĒśäņĀĆĒĢśĻ▓ī ļåÆņØĆ ņäĖĒżļÅģņä▒ņØ┤ Ļ┤Ćņ░░ļÉśņŚłļŗż(Fig. 6).
Ļ│Ā ņ░░
Ļ░£ ņ£ĀņäĀņóģņ¢æņØĆ ļīĆļץ 8ņäĖ ņØ┤ĒøäņØś ņĢöņ╗Ę ļģĖļĀ╣Ļ▓¼ņŚÉņä£ ņŻ╝ļĪ£ ļ░£ļ│æĒĢśļ®░ ļīĆļČĆļČä ņĀäņØ┤ņä▒ņØä Ļ░¢Ļ│Ā ņ׳ļŖö ņĢģņä▒ņØ┤ļŗż. ņ£ĀņäĀņóģņ¢æņØś ņ£ĀĒśĢņ£╝ļĪ£ ņäĀņĢö(adenocarcinoma), ņ£ĀļæÉņĢö(papillary carcinoma), Ļ│ĀĒśĢņĢö(solid carcinoma), ņĢöņ£Īņóģ(carcinosarcoma) ļ░Å ļ│ĄĒĢ®ņĢö(complex carcinoma) ļō▒ņØ┤ ņ׳ļŗż. ļÅÖņØ╝ĒĢ£ Ļ░£ņ▓┤ņŚÉņä£ 1Ļ░£ ņØ┤ņāüņØś ņóģņ¢æ ņ£ĀĒśĢņØ┤ ļ░£Ļ▓¼ļÉśĻĖ░ļÅä ĒĢśļ®░ Ļ░üĻ░üņØś ņóģņ¢æņØĆ ļŗżļźĖ ņóģņ¢æ ļō▒ĻĖēņØä ļéśĒāĆļé┤ĻĖ░ļÅä ĒĢ£ļŗż[12]. ņØ┤ņ▓śļ¤╝ Ļ░£ ņ£ĀņäĀņóģņ¢æņØĆ ņ╣śļŻīĻ░Ć ņēĮņ¦Ć ņĢŖĻ│Ā ņ×¼ļ░£ Ļ░ĆļŖźņä▒ņØ┤ ļåÆņØĆ ņĢöņØ┤ļ»ĆļĪ£, ņóģņ¢æ ņ£ĀĒśĢņŚÉ ļö░ļØ╝ ņ╣śļŻī Ēæ£ņĀüņ£╝ļĪ£ ņé¼ņÜ®ļÉĀ ņłś ņ׳ļŖö ņśłĒøä ņØĖņ×ÉļōżņØä ņĀ£ļīĆļĪ£ ĒÖĢņØĖĒĢśļŖö Ļ▓āņØĆ ļ¦żņÜ░ ņżæņÜöĒĢ£ ņØ╝ņØ┤ļŗż.
Ļ░£ ņ£ĀņäĀņóģņ¢æņŚÉņä£ ņŖżĒģīļĪ£ņØ┤ļō£ ĒśĖļź┤ļ¬¼ ņłśņÜ®ņ▓┤ņØś ļ░£ĒśäĻ│╝ ĻĘĖ ļ░£Ēśä ņĀĢļÅäļŖö ņ¦łļ│æņØś ņ¦äļŗ© ļ░Å ņ╣śļŻīļź╝ ņ£äĒĢ£ ņ£ĀņÜ®ĒĢ£ ņśłņĖĪ ņØĖņ×ÉļĪ£ņä£ ņé¼ņÜ®ļÉĀ ņłś ņ׳ļŖöļŹ░, PRĻ│╝ Ļ░ÖņØĆ ņŖżĒģīļĪ£ņØ┤ļō£ ĒśĖļź┤ļ¬¼ ņłśņÜ®ņ▓┤ņØś ļ░£ĒśäņØ┤ ļČĆņĪ▒ĒĢ£ Ļ▓ĮņÜ░ ņ╣śļŻīņĀü ņĖĪļ®┤ņŚÉņä£ ļ│╝ ļĢī ņóŗņ¦Ć ņĢŖņØĆ ņśłĒøäļĪ£ ņØ┤ņ¢┤ņ¦ł ņłś ņ׳ļŗż. ņĀĢņāü ņ£ĀņäĀņĪ░ņ¦ü ļ░Å ņ¢æņä▒ ņóģņ¢æņĪ░ņ¦üņŚÉņä£ļŖö ņŖżĒģīļĪ£ņØ┤ļō£ ĒśĖļź┤ļ¬¼ ņłśņÜ®ņ▓┤Ļ░Ć ļ░£ĒśäĒĢśņ¦Ćļ¦ī ņĢģņä▒ ņóģņ¢æņĪ░ņ¦ü ļ░Å ņĀäņØ┤Ļ░Ć ņ¦äĒ¢ēļÉ£ ņóģņ¢æņĪ░ņ¦üņŚÉņä£ļŖö PR ņłśņÜ®ņ▓┤Ļ░Ć ņØīņä▒ņØĖ Ļ▓ĮĒ¢źņØä ļéśĒāĆļéĖļŗż[15]. PRņØĆ ņä▒ņן ĒśĖļź┤ļ¬¼ ņāØņé░ņŚÉļÅä Ļ┤ĆņŚ¼ĒĢśņ¦Ćļ¦ī ņóģņ¢æņäĖĒżņØś ņ”ØņŗØņØä ņ£ĀļÅäĒĢśļŖö ņŚŁĒĢĀļÅä ĒĢ£ļŗż[16]. ņØ┤ļ¤¼ĒĢ£ PRņØś ļ░£ĒśäņØĆ ņäĖĒż ņ”ØņŗØ Ļ┤ĆļĀ© ņØĖņ×ÉņØĖ PCNA, Ki67ņØś ļ░£ĒśäĻ│╝ ļ░ĆņĀæĒĢ£ ņāüĻ┤ĆĻ┤ĆĻ│äĻ░Ć ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉśņŚłļŗż. Fanelli ļō▒ņØĆ ņé¼ļ×ī ņ£Āļ░®ņĢö 17Ļ░£ ņĪ░ņ¦üņŚÉņä£ PRņØś ļ░£ĒśäĻ│╝ ņäĖĒż ņ”ØņŗØ Ļ┤ĆļĀ©ņä▒ņØä ņĪ░ņé¼ĒĢśņśĆļŖöļŹ░, PRņØä ļ░£ĒśäĒĢśļŖö 44%Ļ░Ć ņäĖĒż ņ”ØņŗØĻ│╝ Ļ┤ĆļĀ©ļÉśņ¢┤ ņ׳Ļ│Ā 33%ļŖö Ļ┤ĆļĀ©ņØ┤ ņŚåņŚłņ£╝ļ®░ 22%ļŖö PRņØä ļ░£ĒśäĒĢśļŖö ņĪ░ņ¦üņŚÉņä£ ņśżĒ׳ļĀż ļé«ņØĆ ņäĖĒż ņ”ØņŗØņØä ļéśĒāĆļāłļŗżĻ│Ā ļ│┤Ļ│ĀĒĢśņśĆļŗż[17]. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö ļīĆĒæ£ņĀüņØĖ Ļ░£ ņ£ĀņäĀņóģņ¢æņäĖĒżņØĖ REM134ņÖĆ CF41.Mgļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņóģņ¢æ ĒśĢņä▒, ņ”ØņŗØ, ņĀäņØ┤ ļō▒ Ļ┤ĆļĀ© ĒŖ╣ņä▒ņØä ļ╣äĻĄÉĒĢśņśĆļŗż. ĻĘĖ Ļ▓░Ļ│╝, REM134ņŚÉ ļ╣äĒĢ┤ CF41.MgļŖö PR ņłśņÜ®ņ▓┤ņØś ļ░£ĒśäņØ┤ ĒśäņĀĆĒ׳ ļé«ņĢśņ£╝ļ®░, PCNA ļ░Å Ki67ņØś ļ░£ĒśäļÅä Ļ░ÖņØĆ Ļ▓ĮĒ¢źņØä ļéśĒāĆļé┤ņŚłļŗż.
CD133, MDR, Bmi-1 ļ░Å c-mycņØĆ ņóģņ¢æņØä ņ£Āļ░£ĒĢĀ ņłś ņ׳ļŖö ņ×ÉĻ░Ć ņ×¼ņāØļŖźņØä Ļ░Ćņ¦ä ņĢģņä▒ ņóģņ¢æņØś ļīĆĒæ£ņĀüņØĖ ļ¦łņ╗żļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[18]. ĒŖ╣Ē׳, CD133ņØĆ ņżäĻĖ░ņäĖĒż ņåŹņä▒Ļ│╝ ļ¦łņ░¼Ļ░Ćņ¦ĆļĪ£ ņ×ÉĻ░Ć ņ×¼ņāØļŖź, ļČäĒÖöļŖź ļ░Å ļåÆņØĆ ņ”ØņŗØļŖźņØä Ļ░Ćņ¦ä ņäĖĒżņŚÉņä£ ļ░£ĒśäļÉśĻ│Ā ņ׳ņ£╝ļ®░, ņé¼ļ×ī ņ£Āļ░®ņĢö ņ╣śļŻī ņÜöļ▓ĢņŚÉņä£ CD133ņØä ļ░£ĒśäĒĢśļŖö ņäĖĒżļōżņØä Ēæ£ņĀüņ£╝ļĪ£ ĒĢśļŖö ņĀäļץļōżņØ┤ ņĀ£ņĢłļÉśĻ│Ā ņ׳ļŗż[19]. ņĢģņä▒ ņóģņ¢æņØĆ ļģĖĒÖöņŚÉ ņØśĒĢ£ ņäĖĒż ņé¼ļ®Ė ĒÜīļĪ£ļź╝ ĒÜīĒö╝ĒĢśņŚ¼ ļ╣äņĀĢņāüņĀüņ£╝ļĪ£ ņä▒ņןĒĢśĻ▓ī ļÉśļŖöļŹ░, p16INK4a, p53, RB ļśÉļŖö P21WAF1Ļ│╝ Ļ░ÖņØĆ ņäĖĒżņŻ╝ĻĖ░ ņĪ░ņĀł ņ£ĀņĀäņ×ÉņØś ĻĖ░ļŖźņØ┤ ļ╣äĒÖ£ņä▒ĒÖöļÉśĻ▒░ļéś ļÅīņŚ░ļ│ĆņØ┤ļź╝ ņØ╝ņ£╝ĒéżļŖö ĒŖ╣ņ¦ĢņØ┤ ņ׳ļŗż. ņäĖĒżņŻ╝ĻĖ░ ņĪ░ņĀł ņ£ĀņĀäņ×ÉņØś ĻĖ░ļŖźņĀü ļ╣äĒÖ£ņä▒ĒÖöļŖö Bmi-1 ļ░Å c-mycĻ│╝ Ļ░ÖņØĆ ņóģņ¢æ ĒśĢņä▒ ļ¦łņ╗żļź╝ Ļ│╝ļ░£Ēśäņŗ£ĒéżĻ│Ā ņĢöņØś ņ¦äĒ¢ēņØä Ļ░ĆņåŹĒÖöņŗ£ĒéżĻ▓ī ļÉ£ļŗż[18]. Jacobs ļō▒ņØĆ mycņØś ļ╣äņĀĢņāüņĀüņØĖ ņŗĀĒśĖņŚÉ ņØśĒĢ┤ ļéśĒāĆļéśļŖö ņäĖĒżņŻ╝ĻĖ░ ņĪ░ņĀł ņ£ĀņĀäņ×ÉņØĖ INK4a/ARņØś ļ░£ĒśäņØä Bmi-1Ļ░Ć ĒĢśĒ¢ź ņĪ░ņĀłĒĢśņŚ¼ Bmi-1Ļ│╝ c-mycņØś ĒśæļĀź ņ×æņÜ®ņ£╝ļĪ£ ņóģņ¢æ ļ░£ņāØņØä ņ£ĀļÅäĒĢ£ļŗżĻ│Ā ļ│┤Ļ│ĀĒĢ£ ļ░ö ņ׳ļŗż[20]. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö REM134ņÖĆ CF41.Mgļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņĢģņä▒ ņóģņ¢æ Ļ┤ĆļĀ© ĒŖ╣ņØ┤ļ¦łņ╗żļōżņØś ļ░£ĒśäņØä ļ╣äĻĄÉĒĢ£ Ļ▓░Ļ│╝, REM134ņØĆ CF41.MgņŚÉ ļ╣äĒĢ┤ MDR, Bmi-1 ļ░Å c-mycņØś ļåÆņØĆ ļ░£ĒśäņØä ĒÖĢņØĖĒĢśņśĆļŗż.
E-cadherin ļ░Å N-cadherinņØĆ ņ╣╝ņŖś ņØśņĪ┤ņä▒ ņäĖĒż ņĀæņ░®ņØĖņ×ÉļĪ£ņä£ ņäĖĒżĻ░ä ļČĆņ░®ņØä ņ£ĀļÅäĒĢśĻ│Ā ņäĖĒżņØś ņØ┤ļÅÖ ļ░Å ņóģņ¢æņØś ņĀäņØ┤ņŚÉ Ļ┤ĆņŚ¼ĒĢ£ļŗż. E-cadherinņØś ļ░£Ēśä Ļ░ÉņåīļŖö ņäĖĒżĻ░ä ņ£Āņ░®ņØś Ļ░ÉņåīļĪ£ ņØ┤ņ¢┤ņĀĖ ņäĖĒżņØś ņØ┤ļÅÖņØä ņ£ĀļÅäĒĢśĻ│Ā E-cadherinņØś ļ░£ĒśäņØ┤ ņŚåļŖö ņĀäņØ┤ņä▒ ņóģņ¢æ ņäĖĒżņŚÉņä£ N-cadherinņØĆ ļ░£ĒśäļÉśņ¢┤ ņóģņ¢æ ņäĖĒżņØś ņØ┤ļÅÖ ļ░Å ņ£Āņ░®ņØä ņ£ĀļÅäĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[21]. Norval ļō▒ņØĆ[22] ļÅÖļ¼╝ņŗżĒŚś Ļ▓░Ļ│╝ļź╝ ĒåĄĒĢ┤ REM134ļŖö ņĀäņØ┤ņä▒ņØ┤ ņŚåļŗżĻ│Ā ļ│┤Ļ│ĀĒĢśņśĆĻ│Ā, Leonel ļō▒ņØĆ[23] CF41.MgņŚÉņä£ E-cadherinņØś ļé«ņØĆ ļ░£ĒśäĻ│╝ N-cadherinņØś ļåÆņØĆ ļ░£ĒśäņØä ĒÖĢņØĖĒĢśņŚ¼ ņĀäņØ┤ņä▒ņØ┤ ņ׳ļŗżĻ│Ā ļ│┤Ļ│ĀĒĢ£ ļ░ö ņ׳ļŗż. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö REM134ņÖĆ CF41.Mgļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ E-cadherin ļ░Å N-cadherinņØś ļ░£ĒśäņØä ļ╣äĻĄÉĒĢ£ Ļ▓░Ļ│╝, REM134ņŚÉ ļ╣äĒĢ┤ CF41.MgņŚÉņä£ E-cadherin ļ░£ĒśäņØ┤ ļé«ņØīņØä ĒÖĢņØĖĒĢśņśĆņ£╝ļéś, N-cadherinņØś ļ░£ĒśäņŚÉņä£ļŖö Ēü░ ņ░©ņØ┤Ļ░Ć ņŚåņØīņØä ĒÖĢņØĖĒĢśņśĆļŗż.
HER-2ļŖö ņ£ĀņäĀņóģņ¢æņŚÉņä£ ņóģņ¢æņØś ņä▒ņן, ņāØņĪ┤ ļ░Å ļČäĒÖöļź╝ ņĪ░ņĀłĒĢśļŖö ļ¦żņÜ░ ņżæņÜöĒĢ£ ņØĖņ×ÉļĪ£ Ļ░£ ņ£ĀņäĀņóģņ¢æņØś ņĢĮ 30%Ļ░Ć HER-2ļź╝ ļ░£ĒśäĒĢ£ļŗż[12]. HER-2ļź╝ ļ░£ĒśäĒĢśļŖö ņóģņ¢æ ņĪ░ņ¦üņØ┤ ļ░śļō£ņŗ£ ņĢģņä▒ ņóģņ¢æņØ┤ļØ╝Ļ│Ā ĒĢĀ ņłśļŖö ņŚåņ¦Ćļ¦ī HER-2ņØś Ļ│╝ļ░£ĒśäņØĆ ņóģņóģ ļéśņü£ ņśłĒøäļź╝ ļéśĒāĆļéĖļŗżĻ│Ā ļ│┤Ļ│ĀļÉśņ¢┤ ņ׳ņ£╝ļ®░[12], ņé¼ļ×īņŚÉņä£ļŖö ņØ┤ļ»Ė ņŚ¼ļ¤¼ Ļ░Ćņ¦Ć HER-2 ĒāĆĻ▓¤ ņ╣śļŻīņĀ£(trastuzumab, pertuzumab ļ░Å lapatinib ļō▒)Ļ░Ć Ļ░£ļ░£ļÉśņ¢┤ ņ£Āļ░®ņĢö ņ╣śļŻīņĀ£ļĪ£ ņé¼ņÜ®ļÉśĻ│Ā ņ׳ļŗż[24]. ņØ┤ļ¤¼ĒĢ£ HER-2ņØś Ļ│╝ļ░£ĒśäņØĆ ņóģņ¢æņ¢ĄņĀ£ņØĖņ×ÉņØĖ p53ņØś ļÅīņŚ░ļ│ĆņØ┤ņÖĆ Ļ┤ĆļĀ©ļÉĀ ņłś ņ׳ļŗż[25]. p53ņØĆ ļ╣äņĀĢņāüņĀüņØĖ ņäĖĒżņŚÉņä£ ņäĖĒżņé¼ļ®Ė ĻĖ░ļŖźņØä Ļ░¢ļŖö ĒĢĄ ņĀäņé¼ņØĖņ×ÉļĪ£ņä£ ņé¼ļ×ī ņĢöņØś 50% ņØ┤ņāüņØ┤ p53 ņ£ĀņĀäņ×ÉņØś ĻĖ░ļŖźņØ┤ ļÅīņŚ░ļ│ĆņØ┤ļź╝ Ļ░¢Ļ▒░ļéś ĻĖ░ļŖźņØ┤ ņāüņŗżļÉ£ Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[26]. Rom├Īn-Rosales ļō▒ņØĆ p53ņØś ļÅīņŚ░ļ│ĆņØ┤Ļ░Ć ņØ╝ņ¢┤ļéśņ¦Ć ņĢŖņØĆ ņ£Āļ░®ņĢö ņäĖĒżņŚÉņä£ HER-2ņØś ļé«ņØĆ ļ░£ĒśäņØä ļ│┤Ļ│ĀĒĢ£ ļ░ö ņ׳ļŗż[25]. NKņäĖĒżļŖö NKG2DņÖĆ Ļ░ÖņØĆ ļ”¼Ļ░äļō£ļź╝ ļ░£ĒśäĒĢśņŚ¼ ļ│ĆĒśĢļÉ£ ņäĖĒżļōżņØä ņØĖņŗØĒĢśĻ│Ā ņĀ£Ļ▒░ĒĢśņ¦Ćļ¦ī ņóģņóģ NKņäĖĒżĻ░Ć ņĀ£Ļ▒░ĒĢśņ¦Ć ļ¬╗ĒĢśļŖö ņĢöņäĖĒżļōżņØ┤ ņĪ┤ņ×¼ĒĢśļŖöļŹ░, Textor ļō▒ņØĆ p53ņØä ļ░£ĒśäĒĢśļŖö ņäĖĒżņŚÉņä£ NKņäĖĒżņØś ļåÆņØĆ ĒÖ£ņä▒ĒÖöļź╝ ĒÖĢņØĖĒĢśņśĆņ£╝ļ®░, ņ”ē p53ņØś ĻĖ░ļŖźņØ┤ ļ│┤ņĪ┤ļÉśņ¢┤ ņ׳ņØä ļĢī NKņäĖĒż ĻĖ░ļ░ś ļ®┤ņŚŁ ņÜöļ▓ĢņØ┤ ĒÜ©Ļ│╝ņĀüņ£╝ļĪ£ ņ×æņÜ®ĒĢ£ļŗżĻ│Ā ļ│┤Ļ│ĀĒĢśņśĆļŗż[27]. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ CF41.MgļŖö REM134ņŚÉ ļ╣äĒĢ┤ HER-2ņØś ļ░£ĒśäņØ┤ ļé«Ļ│Ā p53ņØä ļåÆĻ▓ī ļ░£ĒśäĒĢśņśĆļŗż. ļśÉĒĢ£, NKņäĖĒżņØś Ēæ£ņĀüņäĖĒżņŚÉ ļīĆĒĢ£ ņäĖĒż ļÅģņä▒ ļ╣äĻĄÉņŚÉņä£ļÅä NKņäĖĒżņŚÉ ļīĆĒĢśņŚ¼ CF41.MgļŖö REM134ņŚÉ ļ╣äĒĢ┤ ļåÆņØĆ ņäĖĒż ļÅģņä▒ņØä ļéśĒāĆļé┤ņ¢┤ NKņäĖĒżņŚÉ ļ»╝Ļ░ÉĒĢśĻ▓ī ļ░śņØæĒĢ©ņØä ĒÖĢņØĖĒĢśņśĆļŗż. Ēśäņ×¼ REM134ļź╝ ņØ┤ņÜ®ĒĢ£ Ļ░£ ņ£ĀņäĀņóģņ¢æ ņŚ░ĻĄ¼Ļ░Ć ļŗżņłśņØ┤ļéś, NKņäĖĒżņŚÉ ļīĆĒĢ£ ĒĢŁņĢö ļ®┤ņŚŁ ņÜöļ▓Ģ ĻĖ░ņ┤ł ņŚ░ĻĄ¼ņŚÉ ņ׳ņ¢┤ņä£ CF41.Mg ļśÉĒĢ£ Ļ░£ ņ£ĀņäĀņóģņ¢æ ņŚ░ĻĄ¼ņŚÉ ņ£ĀņÜ®ĒĢĀ Ļ▓āņ£╝ļĪ£ ĻĖ░ļīĆļÉ£ļŗż.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print