서 론
Fenbendazole (FBZ)은 시중에 유통되고 있는 동물용 항기생충제이며, 요충증을 포함한 다양한 기생충 감염의 치료에 사용되고 있는 약물이다[1-4]. 최근 FBZ의 항암효과가 발견되어 항암제로서의 사용가능성이 연구되고 있다[5-8]. 현재 알려진 FBZ의 항암작용기전은 암세포의 microtubule 중합을 억제하는 anti-tubulin effect를 통해 세포분열을 억제하여 항암효과를 나타내는 것으로 보고되고 있다.
산소는 체내에 존재하며 반응성이 매우 높은 기체로서, 산화적 대사(oxidative metabolism)에 의해 생성되는 자유 전자(free electron)와 쉽게 반응하며, 세포 내에서 superoxide anion (O2−) hydroxyl radical (·O), H2O2와 같은 활성산소(reactive oxygen species, ROS)를 생성한다[9]. ROS의 생산과 항산화물질 작용의 불균형으로 인해 세포 내 ROS가 증가하게 되면 oxidative stress가 발생하며, ROS는 DNA, 단백질 등과 반응하여 손상시키고 cell death를 유발한다[10]. 반면 항암제에 의한 ROS 생산으로 암세포의 사멸을 유도할 수 있다[11,12].
FBZ는 현재 알려진 anti-tubulin effect 이외에 다른 기전을 통하여 항암작용을 갖는 지에 대해서는 연구가 미흡하다. 이에 본 연구에서는 사람 백혈병 세포의 일종인 HL-60세포에 FBZ를 처리하여 ROS와 연관된 항암효과가 유발되는 지 알아보았다.
재료 및 방법
실험세포와 물질 처리
실험에서 사용한 HL-60 세포는 한국세포주은행(Korean Cell Line Bank, Korea)에서 구입하였으며, 10% fetal bovine serum, 2 mM L-glutamine, 100 U/mL penicillin/streptomycin이 포함된 RPMI 1640 세포배지를 이용해 배양하였다. FBZ (Sigma, USA)는 dimethyl sulfoxide (Sigma)로 녹인 후 적정농도로 희석하여 사용하였다.
대사활성도 측정
HL-60 세포를 96-well culture plate에 1 × 105 cells/mL의 농도로 넣은 후, FBZ를 처리하였다. 5% CO2와 37°C의 조건에서 배양한 후 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT, Sigma) assay를 실시하였다[13]. MTT 시약을 well당 0.5 mg/mL의 농도로 4시간 처리한 후 10% sodium dodesyl sulfate 용액을 100 μL/well씩 2시간 동안 처리하여 살아있는 세포에 의해 생긴 crystal violet을 녹였다. 이후 microplate reader (Thermo Fisher scientific, USA)를 이용하여 570 nm에서 흡광도를 측정하였다. 또한 FBZ가 처리된 HL-60세포에서 ROS에 대한 영향을 확인하기 위해, ROS inhibitor인 N-acetyl-L-cysteine (NAC, Sigma)을 3 mM 농도로 처리한 후 MTT assay로 분석하였다.
유세포 분석
세포의 미토콘드리아 막전위(mitochondrial membrane potential, MMP)와 세포사(cell death), ROS를 측정하기 위해 유세포분석을 실시하였다. 세 가지 분석에서 HL-60세포를 24-well culture plate에 1 × 105 cells/mL의 농도로 넣고 FBZ를 농도 별로 처리하였다. MMP를 측정하기 위해 FACS staining solution으로 세척한 뒤 rhodamine 123 (Sigma) 용액을 1 μg/mL의 농도로 처리하여 상온, 암실의 조건에서 30분간 반응시킨 후 분석하였다. 세포사를 측정하기 위해 annexin V-FITC/propidium iodide (PI) 염색을 실시하였다[14]. Annexin V binding buffer (Thermo Fisher scientific)로 세척한 뒤 annex in V-FITC (BD Bioscience)를 1 μg/mL의 농도로 처리한 후 상온, 암실의 조건에서 30분간 반응시켰다. 유세포분석 전에 PI 용액을 0.25 μg/mL의 농도로 세포용액에 넣은 뒤 분석을 하였다. ROS분석을 위해 인산완충액으로 세척한 HL-60세포에 2',7'-dichlorofluorescin diacetate (DCFDA, Sigma) 용액을 10 mM 농도로 30분간 처리하였다. 유세포분석을 위해 염색한 모든 세포는 CytoFLEX® flow cytometer와 CytEx pert® software를 이용하여 분석하였다.
결 과
FBZ처리에 따른 HL-60세포의 대사활성도 변화
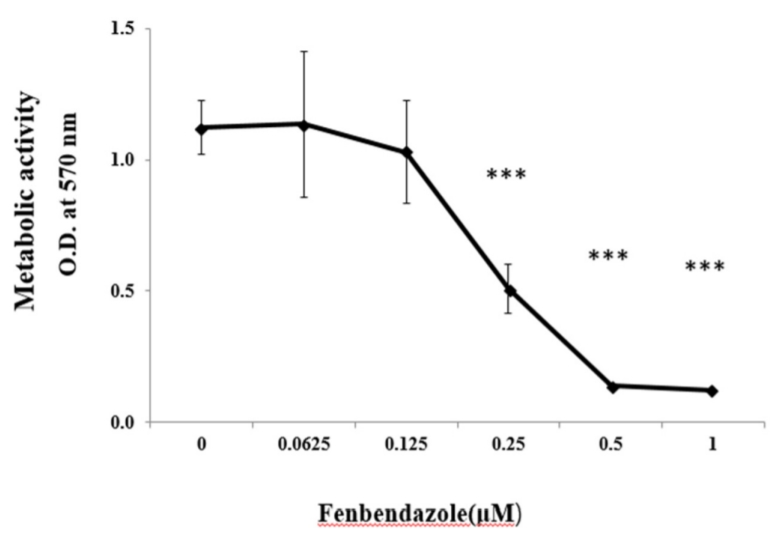
MTT assay는 세포의 대사활성도를 측정할 수 있는 분석 실험이다. MTT 시약의 성분인 tetrazolium이 세포의 대사작용에 의해 보라색의 formazan으로 전환되며 570 nm에서 보라색의 흡광도를 측정함으로써 세포의 대사활성도의 정도를 알 수 있다. HL-60세포에 FBZ를 0-1 μM의 농도로 처리한 결과 0.25-1 μM 처리군에서 세포의 대사활성도가 유의하게 감소하였다(Fig. 1).
FBZ처리에 따른 MMP와 cell death의 변화
FBZ가 처리된 HL-60 세포의 미토콘드리아 막전위 변화를 알아보기 위해 rhodamine 123 염색을 실시한 후 유세포분석을 실시했다. FBZ의 농도 별로 처리된 HL-60 세포를 분석한 결과 처리농도가 증가함에 따라 MMP 측정값은 감소하는 사실을 확인하였다(Fig. 2). MMP는 미토콘드리아의 integrity를 반영하는 지표이며, 감소할 경우 세포의 죽음과 관련이 깊다. 따라서 FBZ에 의해 HL-60세포의 세포사(cell death)가 유도되는 지 알아보기 위해 annexin V-FITC/PI 염색을 한 후 유세포분석을 하였다. FBZ 0.25 μM 처리군은 대조군과 비교하여 차이가 없었지만 FBZ 0.5 μM 처리군에서는 살아있는 세포의 비율이 7%감소하였고, 1 μM 처리군에서는 살아있는 세포의 비율이 30% 이상 감소한 것을 확인했다. 또한 late apoptosis 와 necrosis된 세포의 비율이 대조군에 비해 각각 17%, 18% 증가하였다(Fig. 3).
FBZ처리에 따른 HL-60 세포의 핵 변화
FBZ에 의한 HL-60 세포의 생존율 변화를 시각적으로 확인하기 위하여 세포 핵을 관찰하였다. FBZ가 처리된 세포를 Hoechst 33342 용액으로 염색을 실시한 후 현미경으로 관찰하였다. FBZ 0.25 μM 처리군은 대조군과 비교하여 눈에 띄는 차이가 없었으나 0.5 μM, 1 μM 처리군은 세포의 핵이 깨진 세포들을 확인하였다(Fig. 4).
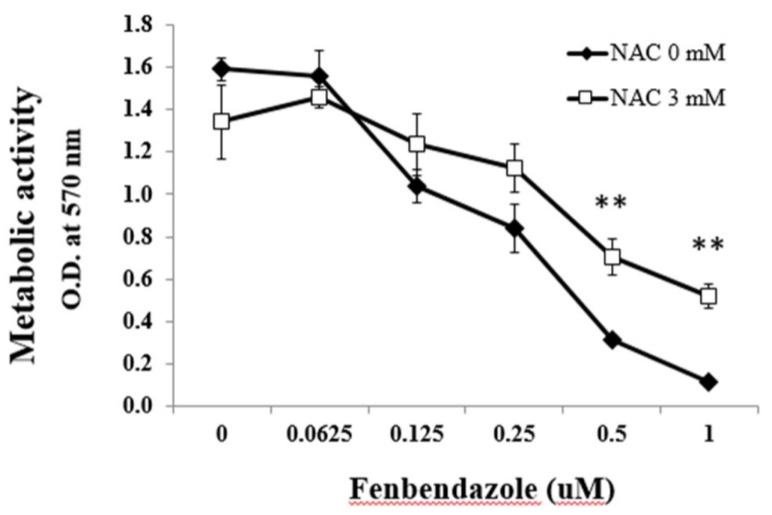
ROS inhibitor 처리에 의한 FBZ의 항암효과 변화
FBZ의 HL-60세포에 대한 항암효과가 ROS생산과 연관이 있는 지 알아보기 위해 세포 내에서 ROS를 감지할 수 있는 시약인 DCFDA를 HL-60세포에 처리한 후 유세포분석을 하였다. 하지만 반복된 실험에도 FBZ에 의한 ROS 생산은 감지할 수 없었다(data not shown). 반면 ROS inhibitor인 NAC를 FBZ와 함께 HL-60세포에 처리하였을 때 FBZ 0.5 μM과 1 μM 농도에서 세포활성도가 유의하게 증가한 것을 확인하였다(Fig. 5).
고 찰
본 연구에서는 동물용 항기생충제이며, 최근 항암효과가 있는 것으로 알려진 FBZ의 항암효과 여부와 ROS 관련 작용기전을 알아보기 위해 연구를 진행하였다. 사람의 백혈병암 세포주(cell line)인 HL-60 세포에 FBZ를 처리한 결과 0.5 μM 이상 처리군에서 미토콘드리아 막전위와 세포활성도가 감소하는 것을 확인하여 FBZ의 항암효과를 확인하였다. 또한 annexin V-FITC/PI 염색을 통해 cell death를 측정한 결과에서도 1 μM 처리군에서 살아있는 세포의 비율이 대조군에 비해 30% 이상 감소하여 FBZ의 항암효과를 재차 확인하였다.
ROS의 생산은 다양한 내부적, 외부적 요인에 의해 시작된다[15]. 암세포는 미토콘드리아의 산화대사가 결핍되어 있어 정상세포에 비해 ROS수준이 높아져 있다[16]. 세포에서의 ROS는 단백질, 지질, 유전자에 영향을 미치므로, 높아진 ROS 수준은 암세포의 성장촉진에 관여한다. 특히 DNA 손상에 의해 종양유전자의 활성과 종양억제유전자의 억제로 발암을 촉진할 수 있다. 그와 반면에 ROS는 간암세포에서 Pglycoprotein (P-gp)의 발현을 촉진시켜 항암제에 대한 저항성을 증가시킨다[17,18]. 실제 항암제 투여 시 폐암과 유방암 세포에서 ROS 생산을 증가시켜 암세포의 세포자멸사를 촉진시키는 사실이 확인되었다[19,20].
ROS에 의한 영향을 확인하기 위해 ROS inhibitor의 일종인 NAC를 사용하였다[21]. FBZ와 NAC를 세포에 처리한 뒤 세포활성도를 측정하였다. 측정 결과 FBZ 0.5 μM 농도부터 NAC 처리군의 세포활성도가 유의하게 증가하였다. 이러한 결과를 통해 FBZ에 의한 항암효과가 일정부분 ROS에 의한 것임을 알 수 있었다. 특히 FBZ의 0.5-1 μM 농도에서 ROS inhibitor인 NAC에 의한 세포활성도 증가는 특정 농도의 FBZ 항암효과가 ROS 생산에 의존하는 것으로 보인다. 하지만 DCFDA를 이용한 직접적인 세포 내 ROS를 측정하지 못했는데, 이는 FBZ를 세포에 처리한 후 ROS의 생산이 급격하게 증가했다가 소멸하는 것이 아닌가 추측한다. 따라서 후속실험을 통해 ROS가 생성되는 시간대를 다르게 하여 추가 실험을 하거나, ROS에 의한 단백질 또는 지질의 손상 정도를 측정하는 분석을 시도할 필요가 있다.
임상에서 많이 사용되는 항암제 중, 미세소관의 형성을 억제하여 암세포의 성장을 막는 항암제에는 paclitaxel, vincristine, vinblastine등이 있다. HL-60 세포를 이용한 in vitro실험에서 paclitaxel은 3.0 nM, vincristine은 4.1 nM, vinblastine은 5.3 nM의 IC50를 보여 강력한 항암효과를 나타냈다[22,23]. 이번 연구에서 FBZ의 IC50는 0.23 μM이었다(Fig. 1). FBZ은 다른 항암제에 비해 약한 항암효과를 보여 주고 있지만, 상대적으로 부작용이 적은 것을 감안하면 단독 투여 또는 기존의 항암제와의 병용투여에 대한 심화된 연구가 필요하다.
본 연구를 통해 항기생충제의 일종인 fenbendazole이 항암 효과를 가지고 있으며, ROS가 작용기전의 일부인 것을 확인했다. Fenbendazole이 현재 항기생충제로 사용되고 있어 항암제로서의 용도를 추가하는 것은 신약개발에 비해 상대적으로 용이할 것으로 판단된다. 이번 연구를 통해 얻은 fenbendazole에 대한 정보는 기초과학 또는 임상 분야에서 유용하게 사용될 수 있을 것으로 사료된다.



 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print