Introduction
Zika virus (ZIKV) is a mosquito-borne virus that was first isolated in 1947 from a febrile sentinel rhesus monkey in the Zika Forest in Uganda [1]. ZIKV belongs to the Flavivirus genus that is classified in the Flaviviridae family along with dengue virus (DENV), yellow fever virus, West Nile virus, and Japanese encephalitis virus. A single-strand positive-sense RNA virus, ZIKV, is composed of a 5′-UTR (~107 nucleotides), a 10.2 kb open reading frame (ORF), and a 3′-UTR (~428 nucleotides), giving a total genome of 11 kb. The ORF encodes three structural proteins (C, prM, and E) and seven nonstructural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) [2,3].
In the past, ZIKV infection was not regarded as a major threat to public health because it induced only mild clinical symptoms such as rash, fever, and headache [4]. However, in 2013, ZIKV began to attract attention as a massive outbreak occurred in French Polynesia [5,6]. In 2015, cases of severe ZIKV infection were reported in Brazil and other South American countries inducing Guillain-Barré syndrome and microcephaly in newborns [7-10]. In addition, ZIKV detection in nonserum body fluids such as semen, saliva, and urine suggests direct human-to-human infection [11,12]. As of mid-2019, ZIKV infection had spread to 87 countries and the Aedes aegypti vector detected in 61 countries. Despite the global spread of ZIKV, no effective vaccine has yet been developed [13].
ZIKV is classified into two main lineages, Asian or African, based on the phylogenetic classification of ORF sequences [14-16]. After the initial detection of ZIKV in Africa, Asian strains spread rapidly to the Pacific islands and the American continent in 2015 and 2016 [4,12,17-19]. The infectivity and pathogenicity of ZIKV are not understood fully, but studies suggest that the African-lineage strain tends to induce more fetal loss and fewer birth defects than the Asian-lineage strain [20,21]. At the amino acid level, the two lineages show differences in 75 to 100 residues of the 3,424 amino acid sequence. This difference between amino acid sequences makes a difference in virus phenotype. For example, SV0127-14 strain, which originated in Thailand, differs by five amino acids from the CPC-0740 strain originating in the Philippines; this difference results in phenotypic differences both in vitro and in vivo [22]. It is possible that susceptibility to ZIKV can be determined by the original mosquitoes vector (Aedes aegypti and Aedes albopictus): studies have shown that ZIKV transmitted via Brazilian mosquitoes was less infectious than strains transmitted by vectors in the Dominican Republic and the United States, which suggests that the virulence might be controlled by viral adaptation to the host [23,24].
Because ZIKV does not replicate efficiently in the presence of type I interferon (IFN), various ZIKV profiling studies have used immunocompromised mouse models such as A129 (type I IFN receptor knockout [KO]), interferon regulatory factor (IRF)3/5/7 triple KO, or AG129 (type I and II IFN KO) [25]. These mice models provide useful data, but these mice strains are limited in use compared to the wild type mice strains because they are expensive. Therefore, in our study, we utilized neonatal and adult immunocompetent mice to develop a mouse model which is less expensive and broad strange of application. And using these mouse models, we compared the virulence of Asian-lineage and African-lineage strain of ZIKV.
Materials and Methods
Mice
Balb/c (female, 8 weeks old) and C57BL/6 (female, 8 weeks old) mice were purchased from Orient Bio Inc. (Korea). Neonatal Balb/c and C57BL/6 mice (one day old) were bred from homozygous breeding pairs. Mice were housed in the Animal Laboratory Center of Kangwon National University under a 12-hour light-dark cycle and given free access to food and water. This work was approved by Institutional Animal Care and Use Committee of Kangwon National University (approval number: KW-180319-1).
ZIKV strains and cell culture
The following ZIKV strains provided by BEI Resources (Manassas, VA, USA) were used in this study: MR766 (BEI Resources No. NR-50065), PRVABC59 (BEI Resources No. NR-50240), and MEX2-81 (BEI Resources No. NR-50280) (Table 1). Sequences of these ZIKV strains were downloaded from NCBI PubMed and aligned using Basic Local Alignment Search Tool (BLAST) (Table 2). Vero and Vero 76 cell lines were purchased from the American Type Culture Collection (Bethesda, MD, USA). Vero cells were incubated in 5% CO2 at 37‚ĄÉ using Minimum Essential Medium (MEM-őĪ) (Gibco, USA) containing 10% fetal bovine serum (FBS) (GenDEPOT, USA) and Vero 76 cells (ATCC, CRL-1587) were incubated in 5% CO2 at 37‚ĄÉ using Dulbecco‚Äôs Minimal Essential Medium (DMEM) (GenDEPOT) containing 10% FBS, until they formed monolayers. Vero cells were infected with PRVABC59 and MEX2-81 strain at a multiplicity of infection (MOI) of 0.01, and then incubated in MEM-őĪ containing 2% FBS. Vero 76 cells were infected with MR766 strain at an MOI of 0.01 and incubated in DMEM containing 2% FBS. Viral samples were concentrated by ultrafiltration and purified using a Capto‚ĄĘ Core 700 column (GE Healthcare, Sweden). Titers of virus stocks were quantified by plaque assay of both cell lines.
ZIKV infection in mice
Eight-week-old Balb/c female mice were inoculated subcutaneously with each strain of ZIKV (MR766 or MEX2-81), or phosphate-buffered saline (PBS) as a negative control. Eight-week-old Balb/c female mice were inoculated subcutaneously with two strains of ZIKV (106.8 TCID50/mouse of MR766 or 106 TCID50/mouse of MEX2-81) or PBS as a negative control. One-day-old Balb/c and C57BL/6 mice were inoculated subcutaneously with each strain of ZIKV (106.8 TCID50/mouse of MR766, 106 TCID50/mouse of MEX2-81, or 106.3 TCID50/mouse of PRVABC59), or PBS as a negative control. Adult mice were monitored until 7 days post infection (dpi) to assess survival rate, weight loss, and abnormal behavior. At 7 dpi, mice were euthanized with CO2 and the weights of organs including heart, lung, liver, brain, and pancreas were measured. Neonatal mice were monitored until 20 dpi to assess survival rate, weight loss, and abnormal behavior. Animals were allocated a score (0-3) [22] based on the most severe clinical sign observed: normal appearance (0); staggering walk, wide stance, or paralysis of hind leg (1); 25% weight loss or labored breathing (2); death (3).
Plaque-forming unit assay
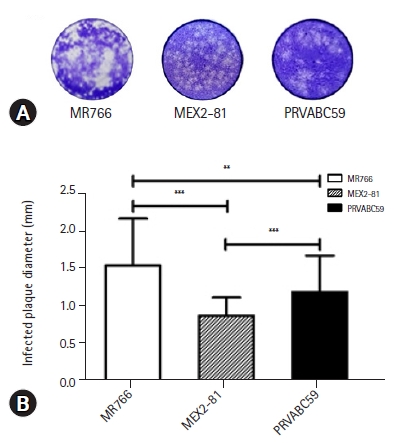
Vero and Vero 76 cells were infected with log dilutions of ZIKV (10-1 to 10-5 dilution) in MEM-őĪ or DMEM containing 10% FBS followed by addition of a 1% agar overlay. After 5 to 7 days, cells were fixed with 8% buffered formalin (in PBS) and stained with 1% crystal violet. Measurements of plaque diameter were made using ImageJ software. Virus titers were calculated by multiplying the plaque count by the virus dilution and dividing by the infection volume, and expressed as plaque-forming units (PFU)/mL. Each sample was assayed in duplicate.
Results
Genetic (phylogenetic) analyses and characterization of ZIKV in vitro
First, we evaluated the virulence of African-lineage strain (MR766) and Asian-lineage strain (MEX2-81, PRVABC59) (Table 1) and compared their nucleotide and amino acid sequences using BLAST alignment (Table 2). For the nucleotide sequence, the African-lineage strain showed approximately 89% similarity to the Asian-lineage strain, whereas the amino acid sequences were approximately 96% identical. As expected, more than 99% similarity was confirmed between the two Asian-lineage strain (Table 2). To examine whether the changes in sequence affected the basic phenotype, we evaluated the characteristics of each strain. When we infected Vero cells with serially diluted ZIKV, the African-lineage strain (MR766) induced a cytopathic effect (CPE) at 4 dpi while that for the Asian-lineage strain (PRVABC59, MEX2-81) CPE occurred 1 to 2 days later. Similarly, the African-lineage strain induced larger plaques (1.6 ¬Ī 0.6 mm) than the Asian-lineage strain (PRVABC59, 1.2 ¬Ī 0.5 mm; MEX2-81, 0.9 ¬Ī 0.2 mm) (Fig. 1). These findings confirmed that the African-lineage strain is more cytopathic than the Asian strains in vitro.
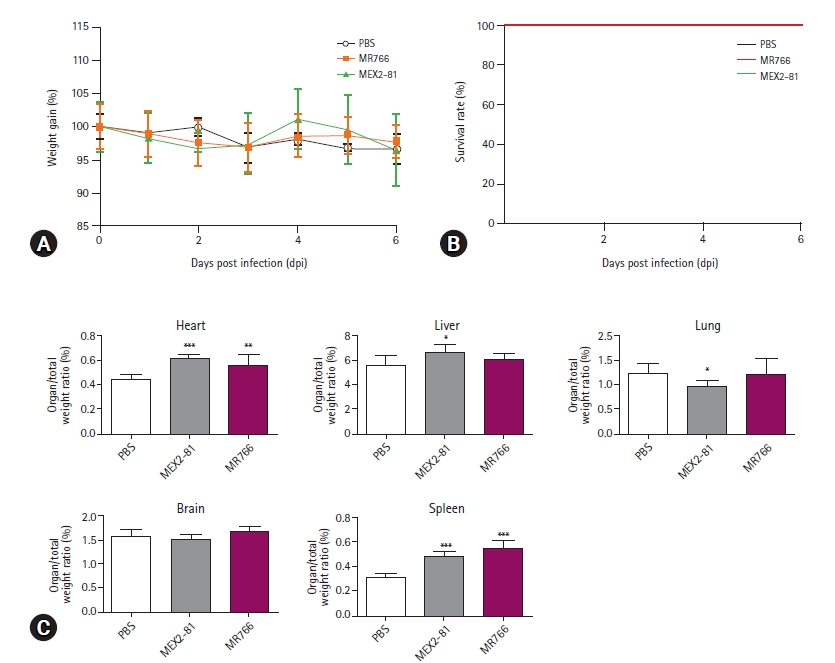
Clinical profile of ZIKV infection in adult mice
Next, we infected 8-week-old female Balb/c to evaluate ZIKA virulence in immunocompetent mice. Infected mice showed no loss in body weight, neurologic signs, pathology, or fatalities (Fig. 2A and B). This is consistent with the findings of previous studies that reported few clinical signs and low susceptibility in immunocompetent hosts. However, we observed a significant increase in spleen weight and size compared to the PBS inoculated group in infected immunocompetent mice, indicating an active immune response to ZIKV infection (Fig. 2C).
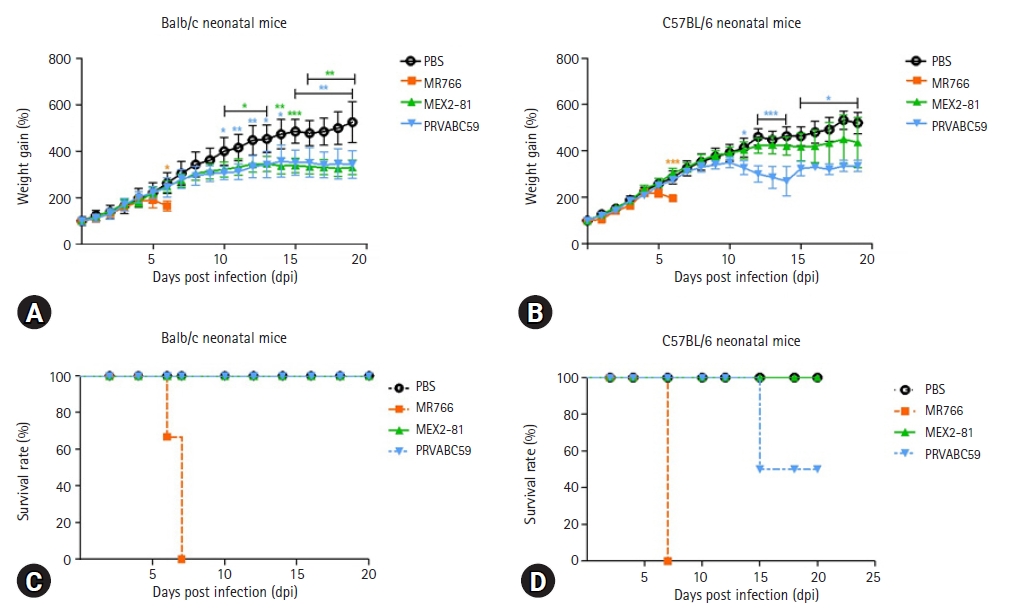
African lineage infection suppressed the growth of neonatal mice
Because the adult immunocompetent mice showed minor clinical signs but indications of an active immune response in lymphoid organs, we attempted to boost the systemic response by infecting neonatal mice in which the immune system had not yet developed. In both Balb/c and C57BL/6 neonates, infection with the African-lineage strain caused a decrease in body weight around 6 dpi and the mice died by 7 dpi. By contrast, Balb/c neonates infected with the Asian-lineage strain showed milder symptoms and all survived up to 20 dpi (Fig. 3A and C). However, C57BL/6 neonates infected with PRVABC59 showed a decrease in body weight from day 5 to day 10 with 40% mortality by 15 dpi; the MEX2-81 strain did not induce any clinical signs by 20 dpi (Fig. 3B and D). These results confirm that the African-lineage strain is more virulent than the Asian-lineage strain, and showed that C57BL/6 neonates were more susceptible than Balb/c neonates, which could be related to their more dominant T helper (Th)1 responses [26].
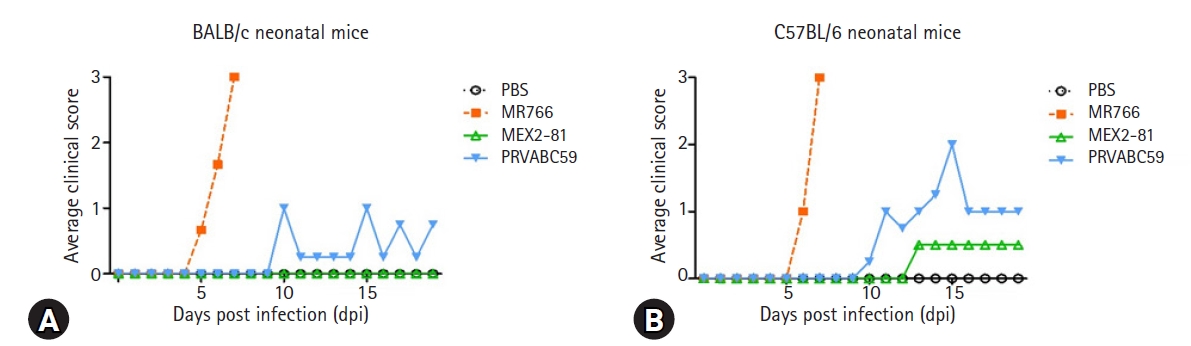
Clinical signs of Asian lineage infection in neonatal mice
Microcephaly is the main pathology induced by ZIKV when it attacks neural progenitor cells. This is associated with various neurological disorders because of abnormal neurogenesis, neuron development, and neural gene expression [27]. We evaluated the neurologic signs in infected mice. Balb/c neonates infected with MEX2-81 strain showed no clinical signs by 20 dpi, but C57BL/6 neonates infected with MEX2-81 strain, half showed neurologic symptoms such as hind leg paralysis by 13 dpi. Balb/c neonates infected with PRVABC59 strain showed symptoms including imbalance and a wide stance at around 10 dpi, but only one of those mice showed paralysis of hind leg after 11 dpi. However, C57BL/6 mice infected with PRVABC59 showed more severe neurologic signs and weight loss from 11 dpi, and 40% of infected neonates died by 15 dpi (Fig. 4). Although the impact of the Asian strain is minimal, our results suggest that Asian strains are also sufficiently virulent to cause significant clinical signs when they infect neonatal mice.
Discussion
Continuous ZIKV outbreaks have been a public health issue for the last several decades, and the possibility of epidemic reemergence of ZIKV requires the development of a vaccine. Although a number of studies have suggested new vaccine strategies, it is still required to develop improved animal models for the accurate evaluation of such vaccines. Thus, our study suggests the possibility of a new, less complicated animal model for ZIKV evaluation. We infected adult and neonatal immunocompetent mice with Asian-lineage and African-lineage strain. Then, we observed their susceptibility and clinical signs in response to infection using different combinations of viral lineage, mouse strain, and mouse age.
In humans, the NS5 protein of ZIKV inhibits type I IFN responses by neutralizing signal transducer and activator of transcription (STAT) 2 signaling. However, this viral protein does not bind to mouse STAT2, thus allowing normal type I IFN responses [28]. Therefore, the majority of ZIKV profiling studies have used type I IFN-deficient mice such as A129 (type I IFNR KO), IRF 3/5/7 triple KO, and AG129 (type I and II IFN KO) [25]. These mice models provide useful data, but these mice strains are expensive and have narrow range of applications due to immunological defects.
The stage of central nervous system development in murine neonates is comparable with that of a human midterm fetus [29]. Furthermore, these neonates are very susceptible to infection with neurotropic viruses including Herpes, Borna virus, Tacaribe arenavirus, and chikungunya virus, which are associated with meningoencephalitis [30-33]. A previous report proposed an animal model to monitor clinical progression, virus distribution, neuropathology, and immune responses after Asian ZIKV infection of C57BL/6 neonates [25]. In the current study, we improved this mouse model by adding the Balb/c mouse strain and using both African-lineage and Asian-lineage strain. It is obvious that type I IFN-deficient mice develop severe clinical signs, but incomplete immunoregulatory mechanisms in these strains may skew our understanding of the pathophysiology of the disease due to impaired IFN response. In addition, previous studies on the pathology of flavivirus suggest that pathogenicity may be determined not only by the effect of the virus but also by the immune response elicited by the virus [32,34]. Therefore, characterizing the clinical changes occurring in immunocompetent hosts will clarify the pathogenic mechanism of ZIKV [26].
Consistent with the findings of previous studies, we observed no significant changes in body weight, survival rates, and neurologic signs when adult mice were infected with various ZIKV strains. However, we observed that the infection resulted in increased weight and size of hearts and spleens. Infection with other arboviruses such as chikungunya virus and DENV induced myocardial effects, and it is possible that ZIKV also invaded cardiac cells [35]. In addition, the increase in splenic size suggests the occurrence of an active immune response in adult mice, even those without clinical signs or changes in body weight. It would be informative to investigate the presence of virus in their peripheral tissues.
Next, we compared the susceptibility of mice to African-lineage and Asian-lineage strain of ZIKV by infecting neonatal mice in which development of the immune system is incomplete. Previous studies reported higher levels of birth defects and fetal loss from infection with African-lineage strain compared with the Asian-lineage strain, which might indicate greater pathogenesis during pregnancy [20,21]. Consistent with this, our results show that the African-lineage strain (MR766) affected all mice but that the Asian-lineage strain (PRVABC59 and MEX2-81) did not affect survival rates, although they induced neurologic symptoms. This means that the African-lineage strain (MR766) is highly virulent and pathogenic strain compared to the Asian-lineage strain (PRVABC59 andMEX2-81) in mice models. Also, our studies are in line with previous observations that ZIKV induces a neurodestructive inflammatory response by activation of cerebral cytotoxic CD8+ T cells [25].
It has been suggested that the amino acid sequence of ZIKV can determine its pathogenicity and virulence. A recent study of the ZIKV/Hu/S36/Ciba/2016 strain reported that the size of plaques was affected by changes in amino acid sequences. For example, viruses with adenine at position 796 (230Gln) formed small plaques but if adenine was replaced with guanine (230Arg), this induced large plaques [36]. The three ZIKV strains used in our study are all 230Arg, but it is possible that the 3.2% to 3.4% variation in amino acid sequence changed viral growth and plaque size. Thus, we speculate that the amino acid modifications were responsible for the differential clinical effects, such as mortality seen with the African-lineage strain and the neurologic signs caused by the Asian-lineage strain. Therefore, a deeper understanding of genetic modulation in ZIKV is essential for vaccine improvement.
In conclusion, our infection model can be applied to investigate the nature of immune responses during ZIKV infection and provide useful information for both vaccine development and selection of challenge strains.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print