Introduction
Pseudomonas aeruginosa is frequently isolated from canines with otitis externa (OE) [1,2]. This pathogen is foreign to the normal otic microflora [3], and many human and mammal studies have provided evidence of biofilm formation by P. aeruginosa [4-8]. A bacterial biofilm is an irreversible structure formed from a bacteria-produced extracellular polymeric matrix. Several steps are involved in the formation of a biofilm. Initially, planktonic bacterial cells attach to a living or nonliving surface and form a stable microcolony. A three-dimensional structure is constructed as the bacterial density increases. The bacterial cells can then detach and transfer to a new site [9,10]. Several studies have reported that bacterial biofilm-formation can be resistant to antibiotic therapy, which results in a persistent and refractory inflammation stage [9-11]. Although the precise mechanisms of biofilm resistance to antimicrobial agents have not been investigated thoroughly, some studies have shown that biofilms inhibit the penetration of antibiotics into bacterial cells, which protect them from the host immune system [9-11]. As a result, antibiotics may be ineffective against bacteria that have formed biofilms. Therefore, some studies have searched for new compounds that may potentiate the efficacy of antibiotics against biofilms.
N-acetylcysteine (NAC) acts as a mucolytic agent and a precursor in glutathione biosynthesis [12-14]. Glutathione is an important modulator for the activity of antibiotics in microorganisms. Hence, NAC has been investigated for its capabilities in inhibiting bacterial biofilms and promoting the antimicrobial activity of known antibiotics [15-20].
This study examined the in vitro efficacy of NAC in combination with antibiotics against the biofilm production of P. aeruginosa. Fourteen P. aeruginosa isolates from canines with OE were tested against NAC with antibiotics commonly used as topical otic treatments for canine OE.
Materials and Methods
Bacterial isolates
Fourteen strains of P. aeruginosa isolates from dogs with OE sampled at animal clinics were used in this study. Clinical samples were cultivated on blood agar and incubated aerobically at 37°C for 24 hours. The isolation of P. aeruginosa was identified through 16S rRNA gene sequencing of the DNA using 27-F and 1492R primers. The National Center for Biotechnology Information database was searched for bacterial DNA nucleotide sequences and compared with the sequences obtained from the isolates.
Determination of minimum inhibitory concentration
NAC (Sigma-Aldrich, USA), enrofloxacin (Sigma-Aldrich), polymyxin B (Sigma-Aldrich), and a gentamicin solution (Sigma-Aldrich) were dissolved in fresh trypticase soy broth (TSB; Kisanbio, Korea) at a starting concentration of 128 μg/mL. The P. aeruginosa inoculate was grown for 24 hours in a 0.5 McFarland suspension in TSB.
The minimum inhibitory concentration (MIC) of NAC, enrofloxacin, polymyxin B, and gentamicin was measured using a broth microdilution assay according to the Clinical Laboratory Standards Institute guidelines. NAC, enrofloxacin, polymyxin B, and gentamicin preparations underwent serial 2-fold microdilution assays. The tested NAC and antimicrobial concentrations were in the range of 0.125 to 64 mg/mL and 0.125 to 64 mg/mL, respectively. A 100 μL P. aeruginosa suspension was added to 100 μL of each working solution in a 96-well microtiter plate (SPL Life Science, Korea). The plate was then incubated in a shaking incubator for 24 hours at 36°C ± 1°C. Positive (only bacterial suspension) and negative (only TSB) controls were also included.
The interactions of NAC + enrofloxacin, NAC + polymyxin B, and NAC + gentamicin were assessed using a 2-fold serial microdilution assay (Fig. 1). The starting NAC concentration was 128 mg/mL, whereas those of enrofloxacin, polymyxin B, and gentamicin were 128 μg/mL. The MICs were determined using the abovementioned method.
The MIC was defined as the lowest concentration that inhibited the growth of bacteria. The fractional inhibitory concentration index (FICI) was calculated to evaluate the potential synergistic or antagonistic activity between NAC and antimicrobials. The FICI was calculated using the following formula: FICA + FICB = FICI, where FICA = MIC of drug A in combination/MIC of drug A alone, and FICB = MIC of drug B in combination/ MIC of drug B alone. The FICI was interpreted as follows: FICI ≤ 0.5 = synergism; 0.5 < FICI ≤ no interaction; FICI > 4.0 = antagonism. All tests were conducted in triplicate.
Biofilm growth
Enrofloxacin, polymyxin B, and gentamicin were dissolved in fresh TSB at a starting concentration of 8,000 ng/mL. The P. aeruginosa isolates were diluted with TSB and standardized to contain 1 × 105 colony forming unit/mL. The enrofloxacin, polymyxin B, and gentamicin preparations underwent a serial 2-fold microdilution assay with concentrations ranging from 7.5 to 4,000 ng/mL. A 100 μL sample of the inoculate suspension was added to each well of a 96-well microtiter plate. The plate was incubated for 12 hours at 35°C to 37°C. The positive (bacterial suspension) and negative (TSB) controls were also included.
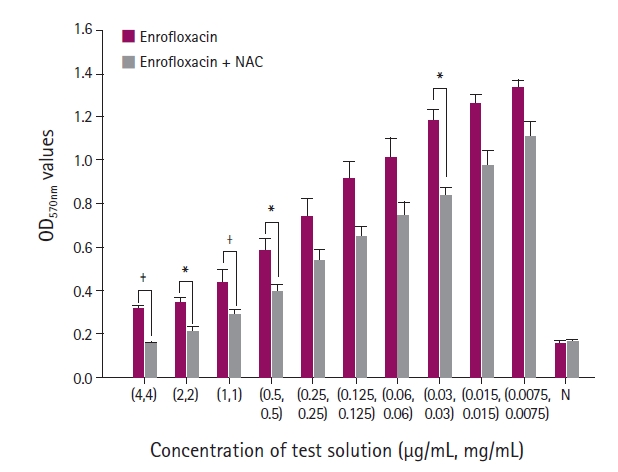
The interaction of NAC + enrofloxacin, NAC + polymyxin B, and NAC + gentamicin were assessed using a 2‒fold serial microdilution assay. The starting concentration of NAC was 4 mg/mL, whereas those of enrofloxacin, polymyxin B, and gentamicin were 4 μg/mL.
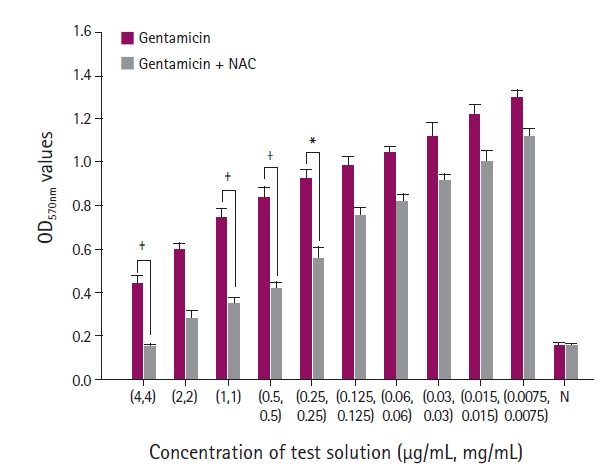
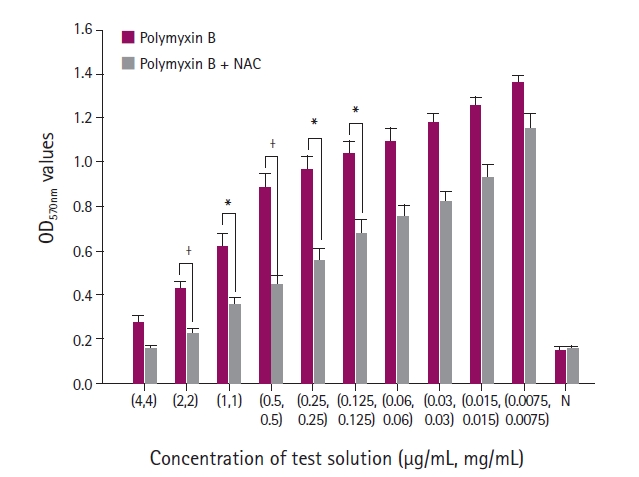
After incubation, the contents in each well were aspirated, and each well was washed three times with 250 μL of phosphate-buffered saline (Sigma-Aldrich). The adherent biofilms were fixed with 200 μL of 99% methanol (Duksan Science, Seoul, Korea) for 15 minutes and stained with 200 μL of 0.1% crystal violet for 5 minutes at room temperature. The excess stain was discarded and rinsed with running distilled water. The microtiter plates were then dried at room temperature for 15 to 30 minutes, and the crystal violet bound to the cells was redissolved with 160 μL of 33% acetic acid (Sigma-Aldrich). The optical density (OD) of each well was measured using a microplate reader (Multiskan Sky; Thermo Scientific, USA) at a wavelength of 570 nm (OD570). The measurements were performed in triplicate and repeated three times. The biofilm production results were divided into four categories based on the criteria reported by Stepanović et al. [21]: no, weak, moderate, and strong biofilm.
Statistical analysis
The results from the triplicate experiments are presented as the mean ± standard error. The differences in the growth of the biofilm exposed to antimicrobial agents with or without NAC were compared using a Mann-Whitney test using SigmaPlot for Windows version 12.0 (Systat Software, USA). p-values < 0.05 were considered significant.
Results
Susceptibility of P. aeruginosa isolates to NAC and antimicrobial agents
Fourteen P. aeruginosa colonies were isolated from canine OE. The MICs of enrofloxacin, polymyxin B, gentamicin, and NAC for P. aeruginosa were 2 to 16 μg/mL, 4 to 32 μg/mL, 2 to 32 μg/mL, and 4 to 8 mg/mL, respectively. The MICs for enrofloxacin, polymyxin B, and gentamicin with NAC for P. aeruginosa were 1 to 4 µg/mL, 1 to 4 µg/mL, and 2 to 4 µg/mL, respectively. Table 1 lists the synergy assays using the MICs.
Interpretation of biofilm production
By applying the criteria of Stepanović et al. [21], P. aeruginosa isolates were categorized into the following group: eight isolates (57.1%) were strong biofilm producers; five isolates (35.7%) were moderate biofilm producers; only one isolate (7.2%) was a weak biofilm producer.
Discussion
P. aeruginosa is a common pathogen that is associated with canine OE [1,2]. It acquires antimicrobial resistance through its ability to form biofilms [6,8,22]. According to previous studies, P. aeruginosa isolated from dogs with OE showed various rates of biofilm production, ranging from 40% to 95% [6-8]. In this study, 14 P. aeruginosa otic isolates with biofilm formation potency were used to evaluate the antibacterial and antibiofilm efficacy of NAC. Before evaluating the abilities of NAC, P. aeruginosa were classified into three groups according to Stepanović et al. [21]: weak, moderate, and strong biofilm producers. In previous reports with the same criteria used in the present study, the distribution across all three groups was similar [6,8]. Pye et al. [6] reported that P. aeruginosa produced 21.2% (7/33) weak, 45.5% (15/33) moderate, and 33.3% (11/33) strong biofilms. Chan et al. [8] showed that the total percentages of weak, moderate, and strong P. aeruginosa biofilms were 31.6% (6/19), 36.8% (7/19), and 31.6% (6/19), respectively. In the present study, more than half of the isolates (57.1%) were strong biofilm producers; only one isolate was a weak biofilm producer. The higher rate of strong or moderate biofilm producers was because P. aeruginosa had been isolated from dogs at a veterinary hospital, as they presented with recurrent or refractory canine OE.
NAC used alone was effective in inhibiting bacteria. Most combinations of NAC with antibiotics displayed different interactions against P. aeruginosa except for four isolates. The rate of synergistic interactions was similar to a previous study, but the rate of antagonistic interactions was lower [20]. Almost all combinations against P. aeruginosa were similar.
The OD values of biofilm stained with crystal violet were measured at 570 nm to evaluate biofilm growth. When the combination of antibiotics and NAC was given to the isolates, the OD values were decreased significantly compared to antibiotics alone. The antibiofilm effect of NAC was maximized dose-dependently.
During biofilm formation, the planktonic bacteria attach to the surface and construct a three-dimensional structure [11,23]. The diffusion of antibiotics into a biofilm becomes challenging once the bacteria form a biofilm from a strong extracellular polymeric matrix. In addition, the biofilm thickness also influences the efficacy of antibiotics [9,10]. Thus, reducing the number of planktonic bacterial and inhibiting thick biofilm formation are key processes for controlling the biofilm resistance to antibiotic agents. Although NAC is usually used as a mucolytic agent in clinics, several studies have shown that this molecule has antibacterial and antibiofilm effects against various microorganisms [5,14,15,17-19]. In the present study, the combination of NAC and antibiotics did not have a significant synergistic effect against P. aeruginosa. On the other hand, NAC alone produced a sufficient decrease in bacterial number. Hence, NAC inhibited biofilm formation when combined with antibiotics.
The antimicrobial and antibiofilm activities of NAC in combination with antimicrobials against P. aeruginosa have been studied [16,17,20]. The present study assessed the in vitro efficacy of NAC combined with enrofloxacin, polymyxin B, and gentamicin against P. aeruginosa isolated from dogs with OE. NAC inhibited planktonic bacterial cells and biofilm formation. The results revealed the efficacy of NAC against the 14 P. aeruginosa isolates evaluated. Almost all strains were strong or moderate biofilm producers. In clinical practice, enrofloxacin, polymyxin B, and gentamicin are antibiotics commonly used as topical treatments for canine OE. Thus, the addition of NAC to common topical solutions for canine OE could be considered a valid and more efficient option against P. aeruginosa otic infections.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print