Expression of the VP2 protein of feline panleukopenia virus in insect cells and use thereof in a hemagglutination inhibition assay
Article information

Abstract
Feline panleukopenia virus (FPV) causes leukopenia and severe hemorrhagic diarrhea, killing 50% of naturally infected cats. Although intact FPV can serve as an antigen in the hemagglutination inhibition (HI) test, an accidental laboratory-mediated infection is concern. A non-infectious diagnostic reagent is required for the HI test. Here, we expressed the viral protein 2 (VP2) gene of the FPV strain currently prevalent in South Korea in a baculovirus expression system; VP2 protein was identified by an indirect immunofluorescence assay, electron microscopy (EM), Western blotting (WB), and a hemagglutination assay (HA). EM showed that the recombinant VP2 protein self-assembled to form virus-like particles. WB revealed that the recombinant VP2 was 65 kDa in size. The HA activity of the recombinant VP2 protein was very high at 1:215. A total of 143 cat serum samples were tested using FPV (HI-FPV test) and the recombinant VP2 protein (HI-VP2 test) as HI antigens. The sensitivity, specificity, and accuracy of the HI-VP2 test were 99.3%, 88.9%, and 99.3%, respectively, compared to the HI-FPV test. The HI-VP2 and HI-FPV results correlated significantly (r = 0.978). Thus, recombinant VP2 can substitute for intact FPV as the serological diagnostic reagent of the HI test for FPV.
Introduction
Feline panleukopenia caused by the feline panleukopenia virus (FPV) and canine parvovirus (CPV) variants 2a, 2b and 2c is the most common and contagious viral disease in cats [1,2]. The clinical signs include vomiting, a high fever, leukopenia, and hemorrhagic diarrhea, resulting in death in about 50% of animals [3]. Since FPV was first identified in cats in the 1920s, it has been isolated from cats, raccoons, monkeys, tigers, and wild carnivores worldwide [4,5]. In South Korea, although many reports have addressed CPV, few have focused on FPV [6,7], which is transmitted via direct or indirect contact with infected animals or contaminated materials. The virus is very stable in the environment, persisting for over 1 year [8].
FPV is a member of the protoparvovirus genus in the family Parvoviridae. The genome is a single-stranded DNA molecule about 5 kb in size that encodes viral proteins 1 and 2 (VP1 and VP2, respectively) and two non-structural proteins (NS1 and NS2) [9-11]. Anti-VP2 antibodies are neutralizing; VP2 contains B-cell epitopes in the N-terminal and loop domains [8,12]. Therefore, the VP2 gene encoding 584 amino acids has been used for evolutionary analysis, and VP2 has been used to detect FPV antibodies after natural infection and vaccination [4,13,14]. VP2 serves as an antigen in the hemagglutination inhibition (HI) test and an FPV subunit vaccine.
Baculovirus expression systems yield high levels of recombinant proteins and virus-like particles (VLPs) [15]. Several recombinant baculoviruses expressing VP2 of CPV and FPV have been created [16,17]. The recombinant VP2 proteins form VLPs via self-assembly and serve as antigens for subunit vaccines, enzyme-linked immunosorbent assays (ELISAs) [17,18], and HI tests.
Virus neutralization (VN), HI, ELISA, and one-step immunochromatography assays have been used to detect anti-FPV and anti-CPV antibodies in animals [19,20]. HI methods are used as standard for detecting such antibodies induced by natural infection or vaccination in dog and cat sera. Although VN tests are accurate and useful for detecting anti-CPV and -FPV antibodies, cell culture and immunostaining are required because FPV does not cause cytopathic effects (CPEs) in small numbers of infected Crandell-Rees feline kidney (CRFK) cells. Thus, VN tests are not appropriate for large-scale sero-surveillance. The HI assay is preferable, because it does not require cell culture. In addition, the HI test is more economical than the VN test and commercial ELISA kits. Currently, no diagnostic reagent for HI of CPV and FPV is available in South Korea. A VP2 protein-based HI test is required for sero-epidemiological studies of FPV.
Here, we expressed recombinant FPV VP2 protein in a baculovirus system and compared the HI titers obtained using FPV and recombinant VP2 to assay cat serum samples.
Materials and Methods
Cells and viruses
CRFK (ATCC CCL-94) cells were grown in Dulbecco’s modified Eagle’s medium with 10% (v/v) heat-inactivated fetal bovine serum (FBS) and an antibiotic-antimycotic cocktail (Gibco, USA). These cells were used to isolate and propagate the FPV1901 strain (KVCC number: VR2100012), which was obtained from a naturally infected Korean cat in 2019. CPV0901 strain (KVCC number: VR0900173) was propagated in A72 cells (ATCC CRL-1542), and used as a control for the detection of optimal hemagglutination assay (HA) activity in various pH condition. Spodoptera frugiperda (Sf9) (CRL-1711; ATCC, USA) cells grown in Grace’s insect medium supplemented with 10% (v/v) FBS were used to propagate the recombinant baculovirus expressing the FPV VP2 protein.
Serum samples
In total, 143 sera from normal cats residing in Chungcheongnam-do and Gyeongsangbuk-do Provinces were collected from 2019 to 2020. All were inactivated at 56°C for 30 minutes prior to the HI assay. We do not know whether the normal cats had been vaccinated against FPV.
Construction of recombinant baculovirus
The full nucleotide sequence of the VP2 gene from FPV1901 was synthesized by Bioneer Co. Ltd. (Korea) with BamH1 and Xho1 restriction enzyme sites at the ends of the open reading frame. A plasmid containing this construct was digested with BamH1 and Xho1. The insert was cloned into the multi-cloning site of the pFastBac HT-B vector to yield FPV-VP2HT, which was transformed into competent Escherichia coli DH10Bac cells (Invitrogen, USA). Recombinant E. coli, identified by polymerase chain reaction (PCR) using the pUC/M13 primer pair (Table 1), was propagated and the recombinant bacmid DNA (BacFPVVP2) was extracted for further transfection. After addition of Sf9 cells to a 24-well plate, mixtures of 1 µg BacFPVVP2 DNA and 8 µL X-treme GENE HP DNA transfection reagent (Roche Diagnostics GmbH, Germany) were added to Sf9 cells according to the manufacturer’s instructions. After incubation for 8 hours at 27℃, the transfection mixture was removed and replaced with new Grace’s medium containing 10% (v/v) FBS. Cells were observed every day and harvested 5 days after transfection. The recombinant V2-expressing baculovirus (ReBac FPVVP2) was passaged three times in Sf9 cells. The third-passage virus served as the viral stock and was propagated to produce recombinant VP2 in Sf9 cells.

The primer sequences used to verify the presence of the feline panleukopenia virus VP2 gene in the recombinant bacmid by polymerase chain reaction
Identification of recombinant VP2 protein using an indirect immunofluorescence assay
An immunofluorescence assay (IFA) was used to identify recombinant VP2 using a mouse anti-CPV monoclonal antibody (APQA, Korea) and a mouse 6x-His-Tag monoclonal antibody (Thermo Fisher Scientific, USA). Sf9 cells infected with ReBac FPVVP2 were fixed in 80% (v/v) cold acetone for 15 minutes and incubated with the two antibodies at 37°C for 1 hour. The cells were washed twice with phosphate-buffered saline (PBS, pH 7.2), stained with fluorescent isothiocyanate-conjugated anti-mouse immunoglobulin (Ig) G + IgM antibodies (200-fold dilution; KPL Laboratories, USA), and examined under a 200× fluorescence microscope.
Identification of VLPs by electron microscopy
Sf9 cells grown in a 150-cm2 flask were infected with ReBac FPVVP2, harvested with 5 mL of culture supernatant 4 days later and subjected to three freeze/thaw cycles. The recombinant VP2 solution was clarified by centrifugation at 3,000 × g for 15 minutes; the supernatant was layered onto a cesium chloride solution with a density 1.43 g/cm2 and centrifuged at 100,000 × g for 2 hours using a SW-41 rotor. The band on top of the cesium chloride layer was collected and injected into a Slide-A-Lyzer dialysis tube (Thermo Fisher Scientific). After dialysis against PBS overnight at 4°C, one drop was placed on a Formvar-coated grid and negatively stained with 1% (w/v) uranyl acetate. ReBac FPVVP2 VLPs were visualized by electron microscopy (EM).
Western blotting of recombinant FPV VP2
The molecular weight of FPV VP2 was determined by Western blotting (WB). The supernatant, cell lysate, and FPV1901 samples were separated on a 12.5% (w/v) Tris-glycine gel and transferred to a nitrocellulose membrane, which was exposed to an anti-CPV VP2 monoclonal antibody (APQA) overnight at room temperature. Secondary antibody (horse radish peroxidase–conjugated goat anti-mouse IgG antibody; 1:2,000 dilution) was then added followed by incubation for 1 hour. After washing, 3ʹ3-diaminabenzidine was added for color development.
HA and HI tests
The optimal HA activities of three antigens (FPV1901, recombinant VP2 protein, and CPV0901) were measured in Sorensen solution at pH 5.8 to 6.8. FPV1901 and recombinant VP 2 protein were then subjected to serial two-fold dilution in 50 µL amounts of buffer at pH 6.0 in a 96-well (U-form) microplate; 50 µL amounts of a 0.6% (w/v) pig erythrocyte suspension with 0.1% (w/v) bovine serum albumin in Sorensen buffer was added and the plate was incubated at 4℃ for 1 hour. The HA titer was the reciprocal of the highest dilution of FPV1901 or recombinant VP2 exhibiting an HA reaction. HI tests were then performed using the two antigens in the microplates. The HI test details have been published previously [20]. The HI titers measured using FPV1901 and recombinant VP2 were compared.
Statistical analysis
Least-squares linear regression analysis was employed to determine the correlation coefficient (r value) between two HI titers. The r value was automatically calculated by Microsoft Excel 2010 software (Microsoft Corp., USA).
Results
Identification of recombinant VP2 protein
The VP2 gene of FPV1901 was cloned into the pFastBac HT-B vector and the recombinant plasmid, FPV-VP2HT, identified by BamH1/Xho1 double digestion. BacFPVVP2 DNA was also PCR-checked using the bacmid primer pair. As shown in Fig. 1, the VP2 genes in the pFastBac HI-B vector and BacFPVVP2 DNA were 1,755 and 4,230 bp in size, respectively. Sf9 cells infected with ReBac FPVVP2 exhibited CPEs, such as rounding and shedding (Fig. 2A); normal Sf9 cells did not show CPEs. Expression of recombinant VP2 in Sf9 cells was verified by IFA; the addition of a monoclonal anti-CPV antibody and anti-6-histidine antibody resulted in obvious green fluorescence (Fig. 2C and D). To visualize the VLPs of ReBac FPVVP2, the virus was propagated in Sf9 cells and VLPs purified by cesium chloride centrifugation. EM revealed that the VLPs were about 22 nm in size, typical of the Parvoviridae (Fig. 3). The cell supernatant and lysate were subjected to WB; the monoclonal anti-CPV antibody identified proteins with a molecular weight of 65 kDa in both the recombinant VP2 preparation and FPV1901 (Fig. 4). The HA activities of FPV1901 and ReBac FPVVP2 were examined at various pH values. The optimal pH of Sorensen buffer, i.e., that providing the highest HA titers, was pH 6.0 (Table 2). The cell supernatant and lysate were used in the HA test; as shown in Fig. 5, the HA titer of the lysate was 1:215.

Identification of the VP2 gene in the pFastBac HT-B vector and the bacmid. Insertion of the VP2 gene into the pFastBac HT-B vector was confirmed by BamH1/Xho1 double digestion. The bacmid containing the VP2 gene was identified by polymerase chain reaction. M, 1-kb DNA ladder, lane 1, pFastFPVVP2 plasmid; lane 2, Bacmid DNA containing the VP2 gene. FPV, feline panleukopenia virus.

Identification of recombinant feline panleukopenia virus (FPV) VP2 baculovirus. The cytopathic effects in Sf9 insect cells infected with recombinant FPV VP2 baculovirus (A); and normal Sf9 insect cells (B). Immunofluorescence of Sf9 insect cells infected with recombinant baculovirus expressing VP2, as revealed by a mouse monoclonal antibody against VP2 (C) and an anti-6-histidine antibody (D).
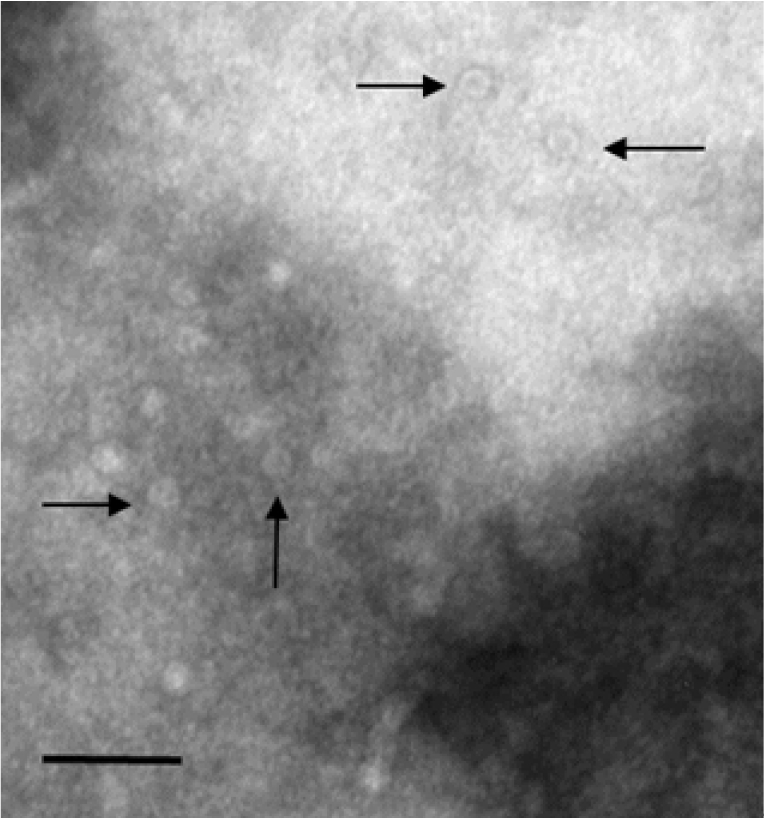
Transmission electron micrograph of ReBac FPVVP2. Scale bar: 100 nm. Arrows indicate the virus-like particles of feline panleukopenia virus. FPV, feline panleukopenia virus.
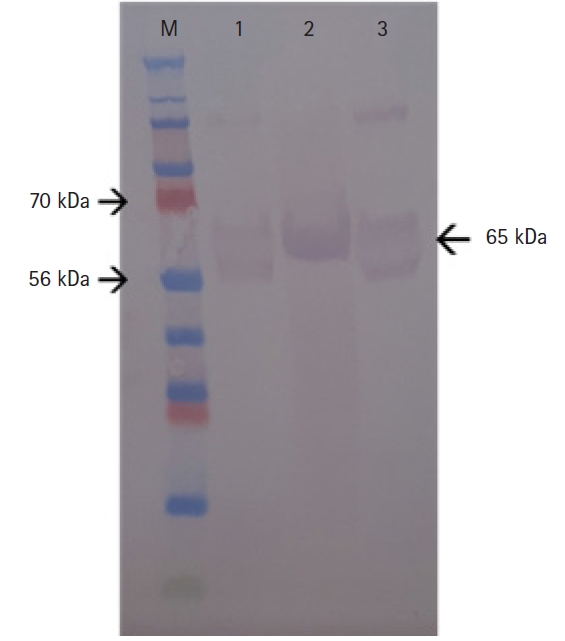
Western blotting of the recombinant feline panleukopenia virus VP2 protein expressed by a recombinant baculovirus using a monoclonal antibody against parvovirus. The molecular weight of recombinant VP2 was 65 kDa. M, protein ladder; lane 1, supernatant; lane 2, cell lysate; lane 3, feline panleukopenia virus.
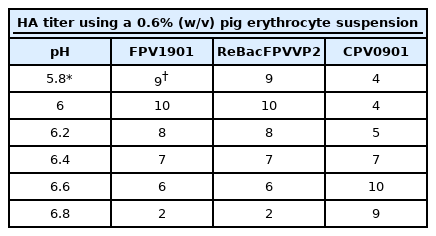
The HA activities of FPV1901 and the recombinant VP2 protein
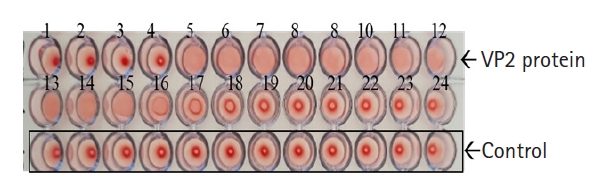
Hemagglutination assay (HA) activity of recombinant VP2 protein of feline panleukopenia virus. The HA titer is 1:215. The phenomenon that looks like a negative reaction in columns 1 to 4 is due to the prozone phenomenon.
Comparison of the HI-FPV and HI-VP2 tests
The two assays were used to detect FPV HI antibodies in 143 cat serum samples. Table 3 shows that 135 and 134 were positive on the HI-FPV test (titer ≥ 1:20) and new HI-VP2 test, respectively. The HI-FPV and HI-VP2 test identified nine and eight FPV-negative samples, respectively. The sensitivity, specificity, and accuracy of the HI-VP2 test were 99.3%, 88.9%, and 99.3%, respectively, compared to those of the HI-FPV test (Table 3). Fig. 6 shows the correlation between the HI-FPV and HI-VP2 data. The r value of the HI-VP2 test was 0.978.

The sensitivity, specificity, and accuracy of the HI-VP2 test for detecting FPV antibodies in comparison with the HI-FPV1901 test
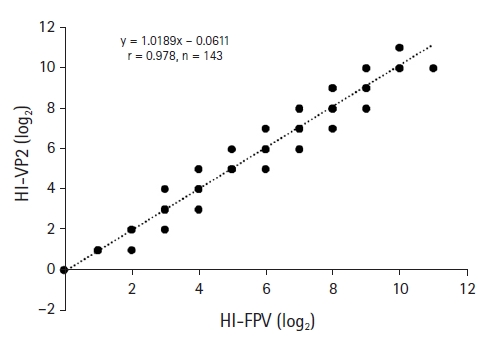
Correlation between feline panleukopenia virus (FPV) antibody titers obtained using the hemagglutination inhibition (HI)-FPV and HI-VP2 tests to evaluate 143 cat serum samples. The correlation is indicated by the linear regression line.
Discussion
Several systems have been used to express VP2 of CPV and FPV. VP2 contains the major viral epitopes. PCR amplification of the VP2 gene has been used to detect wild FPV [21]. VP2 has been expressed in several hosts, including E. coli [22], insect cells [16,23,24], and a plant [25], for use as a subunit vaccine. We expressed VP2 in a baculovirus system, as revealed by IFA, WB, EM, and an HA test. Bac-to-Bac systems expressing foreign proteins have many advantages, such as appropriate post-translation modification and protein processing [23]. A recombinant VP2 protein vaccine is a safe alternative to a whole-virus vaccine; vaccination with live FPV is associated with FPV and CPV DNA in the white blood cells of asymptomatic cats [1]. A VP2 subunit vaccine could replace the traditional live vaccine [24]. Our recombinant VP2 expressed in insect cells may be a suitable replacement.
Serological FPV surveys can determine field infection and vaccination rates. Although the VN test is accurate, many veterinary laboratories prefer to use the simpler and cheaper HI and ELISA tests. Thus, we developed an HI method using recombinant VP2 protein for sero-surveillance of FPV antibodies in normal cats, and compared it to the conventional HI-FPV test. The conventional HI method uses an infectious FPV strain to measure antibodies in sera. The HA activity of FPV propagated in CRFK cells does not exceed 1:210. Our recombinant VP2 protein exhibited an HA activity of over 1:215 and is non-infectious. Thus, our recombinant VP2 can serve as an antigen in the HI test.
The sensitivity, specificity, and accuracy of the HI-VP2 test, and the correlations with the results of the traditional HI test, were determined by analyzing 143 cat serum samples. The sensitivity (99.3%) of the HI-VP2 test for FPV antibody detection was significantly higher than the specificity (88.9%). Only 1 of 135 positive and 8 negative samples showed differences between by the two test methods. One of nine HI-FPV-negative samples was missed by the HI-VP2 test (11.1% false-positive rate). This false positive reaction showing a difference of one level in HI test may reflect a non-specific reaction [26]. It is necessary to test a large number of negative samples to identify the cause of the false-positive.
The HI-VP2 and HI-FPV HI titers were well-correlated (r = 0.978); the HI-VP2 test was thus as reliable as the HI-FPV test. A new I-ELISA using purified recombinant VP2 can readily detect FPV antibodies in cat sera.
In conclusion, we expressed recombinant FPV VP2 using a baculovirus expression system. We detected viral CPEs, performed an IFA and WB, derived HA values and applied EM analysis. The recombinant VP2 protein in a cell lysate had a very high HA titer (1:215) and can thus serve as a new diagnostic reagent for HI tests to detect FPV.
Acknowledgements
This work was supported financially by a grant (B1543083-2020-22-01) from Animal, and Plant Quarantine Agency, Ministry of Agriculture, Food and Rural Affairs (MAFRA), Republic of Korea.
Notes
The authors declare no conflict of interest.