Introduction
The developmental competence of embryos derived from somatic cell nuclear transfer (NT) is lower compared to those from parthenogenetic activation (PA), in vitro fertilization (IVF), or intracytoplasmic sperm injection [1,2]. NT success rate in mammals has not changed significantly since the beginning [3,4]. Many factors are involved in the low pregnancy rate [5]. NT is a complex multistep procedure, and external stress can reduce the quality of NT-derived embryos. During these steps, normal or abnormal reprogramming may occur, which possibly explains many defects occurring in the later reprogramming stage [6]. Low developmental capacity hinders the efficient production of customized embryonic stem cells or animals for organ transplantation and the development of disease models, respectively.
Higher organisms, such as mammals, were considered incapable of parthenogenesis. However, PA technology was recently used to birth a mouse from unfertilized oocytes [7,8]. PA-derived embryos usually get arrested in the preimplantation stage without gene editing. However, PA-derived embryos aggregated with normal embryos can develop into normal chimeric offspring [9]. These results indicate a possible way to produce high-quality early-stage embryos by aggregating low-quality and high-quality PA embryos.
The ability to produce blastocysts and create chimeric animals and embryonic stem cell lines can be improved by producing high-quality blastocysts through blastomere aggregation [10-12]. By addressing developmental issues in vitro, the embryo aggregation approach has been applied to many mammals to increase production efficiency. For example, pigs exhibited improved blastocyst quality and cell viability in response to embryonic aggregation. Blastomere aggregation improves embryonic development by reducing apoptosis and promoting cell reprogramming [13,14].
While the zona pellucida (ZP) is essential in fertilization [15], its removal is necessary for blastomere aggregation, successfully handling early-stage blastomeres for differentiation analysis, producing identical twins or quadruplets, and introducing genes. Tyrode’s solution (TS) or protease (pronase E, PE) are frequently utilized for zona-removal; however, prolonged exposure can hinder development and lead to the collapse of the three-dimensional blastomere structure [15]. Therefore, it is necessary to study the effects of the removal method on developing porcine embryos.
The well-of-the-well (WOW) method is a standard method for embryo aggregation [16,17]. However, in a previous study, we examined the aggregation of PA-derived pig embryos using phytohemagglutinin-L (PHA-L) [12]. The aggregation method of blastomeres using PHA-L produced blastocysts with improved quality and establishment efficiency of embryonic stem-like cells. In addition, the method's safety was confirmed by normal three-germ layer differentiation after induction into embryoid bodies.
Compared to embryos derived from other artificial reproductive technologies, PA embryos are more resistant to external factors and can be produced using simple electrical stimulation. Further, the aggregation of PA-, NT-derived embryos with low embryonic developmental competence could have a synergistic effect. In this study, we established the most efficient method for ZP removal and blastomere aggregation in pigs. We also investigated whether the aggregation of NT and PA blastomeres could reduce embryonic apoptosis and improve the quality of NT-derived embryos.
Materials and Methods
Culture media
All compounds, unless otherwise indicated, were bought from Sigma-Aldrich Corporation (USA). Oocytes were allowed to mature in Medium 199 (M‐199; Invitrogen, USA) supplemented with 0.91 mM pyruvate, 0.6 mM cysteine, 10 ng/mL epidermal growth factor, and 1 µg/mL insulin, with 10% (v/v) porcine follicular fluid as the base in vitro maturation medium. The in vitro culture media was porcine zygote medium 3 supplemented with 0.3% (w/v) bovine serum albumin (BSA). The embryos were moved to drops of porcine zygote medium-3 with 10% fetal bovine serum (ThermoFisher Scientific, USA) on day 4.
Oocyte collection and in vitro maturation
The in vitro maturation of oocytes was performed in the same manner as described in Lee et al. [12]. The ovaries of prepubertal gilts, obtained from a local abattoir, were transported to the laboratory at 34°C to 37°C. Cumulus oocyte complexes (COCs) were aspirated from follicles of 3-8 mm in diameter. COCs with multiple layers of compacted cumulus cells were selected and washed three times in HEPES-buffered Tyrode’s medium (TLH) containing 0.05% (w/v) polyvinyl alcohol (TLH‐PVA). COCs were placed in each well of a four-well multi-dish (Nunc, Denmark). Each well contained 500 µL of in vitro maturation medium with 10 IU/mL equine chronic gonadotropin and 10 IU/mL human chorionic gonadotropin (Intervet, The Netherland). After 22 hours, the COCs were transferred to a fresh in vitro maturation medium, without equine or human chorionic gonadotropin, and incubated for 22 hours. In vitro maturation was performed at 39°C and 5% CO2 in a humid incubator (Astec, Japan). After in vitro maturation, COCs were denuded by gentle pipetting with 0.1% hyaluronidase.
PA and NT
NT and PA were performed as previously described in Lee et al. [18]. For NT, green fluorescent protein (GFP)-inserted porcine fetal fibroblasts were used as NT donor nuclei. Donor cells were cultured in Dulbecco’s modified Eagle medium F-12 (Invitrogen) supplemented with 10% (v/v) fetal bovine serum until a complete monolayer of cells was formed. The donor cells were synchronized at the G0/G1 stage of the cell cycle for 3 to 4 days by contact inhibition. A suspension of single cells was prepared using EDTA-trypsin and resuspending in TLH containing 0.4% (w/v) BSA (TLH-BSA) before cell injection. In vitro matured oocytes were incubated for 10 minutes in TLH-BSA medium containing bisbenzimide H33342, washed twice with fresh TLH-BSA medium, and then transferred to a droplet of TLH-BSA containing 5 μg/mL cytochalasin B covered with mineral oil. The first polar body and adjacent cytoplasm containing the chromosomes were enucleated using a 16-μm polar body biopsy pipette (Sunlight Medical, USA). A donor cell was inserted into the perivitelline space of an enucleated oocyte. The reconstituted single cell-oocyte couplets were simultaneously induced to fuse and activate through a single electrical stimulation. Fusion-activation was induced using an alternating current field of 2 V at 1 MHz for 2 seconds, followed by two pulses of 170 V/mm direct current for 30 μsec using an Electro Cell Fusion Generator (LF101; NepaGene, Japan) in a fusion-activation medium consisting of 280 mM mannitol, 0.05 mM MgCl2, and 0.1 mM CaCl2. Electrically stimulated oocytes were transferred to porcine zygote medium-3 supplemented with 1.9 mM 6-dimethylaminopurine combined with 0.4 μg/mL demecolcine for 4 hours at 39°C temperature and in a humidified atmosphere of 5% CO2, 5% O2, and 90% N2.
Only oocytes from which the first polar body was released after in vitro maturation were selected for PA. They were placed between two wires in a 1 mm fusion chamber coated with an activation medium (280 mM mannitol, 0.05 mM MgCl2, and 0.1 mM CaCl2). Oocyte activation was stimulated with a direct current pulse of 120 V/mm for 60 μs using Cell Fusion Generator. Electrically stimulated oocytes were transferred to porcine zygote medium-3 with 5 μg/mL cytochalasin B and cultured for 4 hours at 39°C temperature in a humidified atmosphere of 5% CO2, 5% O2, and 90% N2.
ZP removal and blastomere aggregation
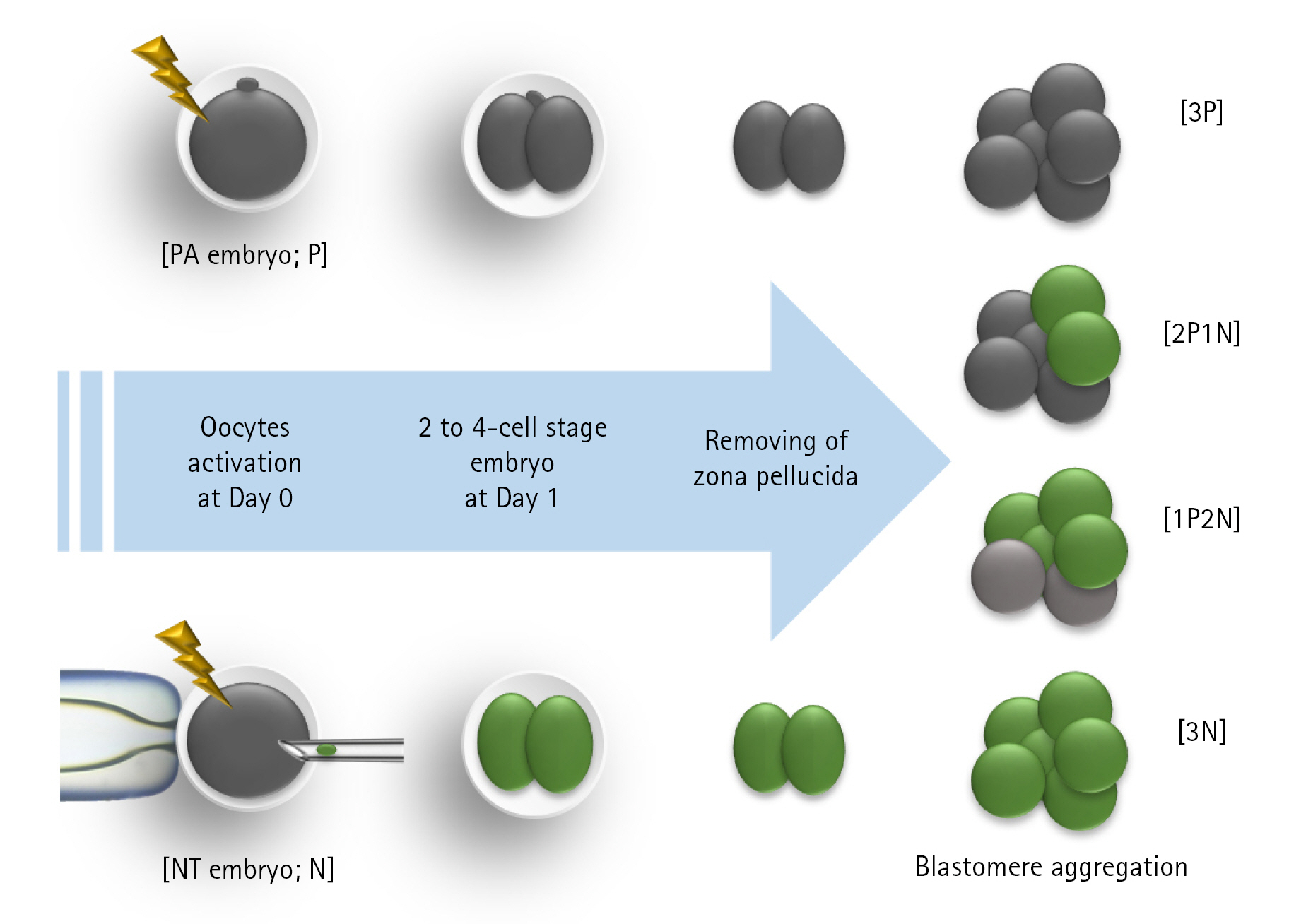
According to the experimental design, the 2 to 4 cell stage PA or NT embryos were collected 24 hours after electrical activation, respectively. The collected embryos were incubated with TS (Gibco, USA) or PE to remove the ZP. ZP removal using PE was performed as previously described by Li et al. [19]. The embryos were placed in 0.3% (w/v) PE for 30 seconds and immediately washed twice or thrice in TLH-BSA. The remaining ZP was removed mechanically using a glass pipette. The ZP was removed using TS as previously explained by Lee et al. [12]. The collected embryos were incubated in 50% TS solution for 1 minute, 100% TS solution for 30 seconds, and then 50% TS solution for 1 minute to remove all of the embryos that needed to be removed. The PHA-L or WOW technique was used to aggregate the ZP-free blastomeres [12]. Blastomeres were aggregated by cultivating the blastomeres for 20 minutes within in vitro culture medium with 15 μg/mL PHA-L. Micro-well dishes were prepared for the WOW system one day before embryo cultivation. To push down on the bottom of a polystyrene dish (Falcon REF351007; Coning, USA), a glass ball with a diameter of roughly 550 to 600 m was heated using an alcohol lamp. The pressure was then maintained until the glass balls were sufficiently cooled (about 5 seconds). A dish with a 60 mm diameter was made with about 30 micro-wells and used.
Experimental design
In Experiment 1, the effects of the ZP removal method on survival, aggregation, and blastocyst development of PA-derived blastomeres were investigated. In Experiment 2, the effects of the aggregation method on blastocyst development and blastocyst diameter of NT-derived blastomeres were investigated. Individually cultured embryos were used as a control, and blastomere aggregation efficiency using WOW and PHA-L was examined. In Experiment 3, the effects of the aggregation of PA-derived blastomeres and NT-derived blastomeres on embryonic development were investigated (Fig. 1). Blastomeres were aggregated from three PA blastomeres (3P), two PA blastomeres with one NT blastomere (2P1N), one PA blastomere with two NT blastomeres (1P2N), and three NT blastomeres (3N). In Experiment 4, we investigated apoptosis in aggregate-derived blastocysts.
Evaluation of developmental competence of blastocyst and total cell count
Day 0 was considered the first day that PA or NT began. Cleavage formation was examined on day 1, and on day 7, blastocyst formation was evaluated. Aggregation efficiency refers to the proportion of aggregates still present in a single lump on day 7 to the total cultured aggregates. Blastocysts were fixed in 4% (v/v) paraformaldehyde in phosphate-buffered saline containing 0.05% polyvinyl alcohol and stained for 5 minutes with 5 μg/mL of Hoechst-33342 to determine the total number of cells present on day 7 of development. Following that, each group's blastocysts were moved to a drop of 100% glycerol on a glass slide and delicately covered with a coverslip. The stained blastocysts were examined using a fluorescent microscope (Nikon, Japan) with a 200X magnification.
TUNEL assay
Apoptosis analysis was carried out, as previously explained by Lee et al. [12]. Blastocysts were fixed for 1 hour at room temperature (RT) with 4% (v/v) paraformaldehyde, rinsed with Dulbecco’s phosphate-buffered saline (DPBS) containing 0.05% polyvinyl alcohol, then permeabilized for 1 hour at RT with 0.1% (v/v) Triton X-100 in 0.1% (w/v) sodium citrate. The embryos were stained with 45 μL of TUNEL-Label solution (Roche, Germany) and 5 μL TUNEL-Enzyme solution (Roche) for 1 hour at 39°C in a dark, humid environment after rinsing with DPBS containing 0.05% polyvinyl alcohol. The nuclei were stained for 10 minutes with 5 μg/mL Hoechst-33342.
Evaluation of GFP expression in the blastocyst
NT donor cells were injected into enucleated oocytes by selecting only cells expressing GFP using an ultraviolet (UV) filter. After 6 days of blastomere aggregation, blastocyst-type embryos were collected, washed three times with TLH-PVA, placed in 10 μL drops of TLH-PVA, and GFP expression was observed using an epifluorescence microscope (TE300; Nikon) equipped with a 460 nm UV filter (Fig. 2).
Statistical analysis
The Statistical Analysis System software was used to conduct statistical analysis (ver. 9.4; SAS Institute, USA). The data were analyzed using a general linear model procedure followed by the least-significant-difference mean separation procedure when there were differences between treatments (p < 0.05). Prior to analysis, percentage data were arcsine-transformed to preserve variance homogeneity. The results are expressed as the mean and standard error of the mean.
Results
Effects of ZP removal method on survival, aggregation, and blastocyst development of PA-derived blastomeres
In the first experiment, the survival rate, aggregation rate, and blastocyst developmental competence of PA-derived blastomere aggregates according to the removal methods of the ZP were investigated. The aggregation rate of embryos treated with PE was significantly (p < 0.05) decreased compared to those of embryos treated with TS (69.2% ± 4.2% vs. 88.3% ± 4.4%). However, the two treatment groups had no significant difference in survival and blastocyst development rate (Table 1).
Effects of aggregation method on blastocyst development and blastocyst diameter of NT-derived blastomeres
In the second experiment, NT-derived blastomeres were aggregated using the WOW or PHA-L methods to investigate the blastocyst development rate and diameter of the blastocysts. No significant difference in the rate of blastocyst development was observed between the PHA-L and WOW methods (35.7%-38.5%). However, the aggregate-derived blastocysts showed significantly increased diameters compared to the control group (155 μm vs. 193-203 μm) (Table 2).
Developmental competence of chimeric embryos produced by NT- and PA-derived blastomeres aggregation
In the third experiment, the 2P1N treatment group showed a significant increase in the rate of blastocysts compared to the 3N treatment group (73.8% ± 9.5% vs. 24.3% ± 10.9%). We found that the diameter of the blastocysts in the 2P1N treatment group was significantly greater than those in the control and 1P2N group (274 ± 12.1 μm vs. 230-234 μm) (Table 3).
Apoptosis of chimeric blastocysts produced by the aggregation of NT- and PA-derived blastomeres
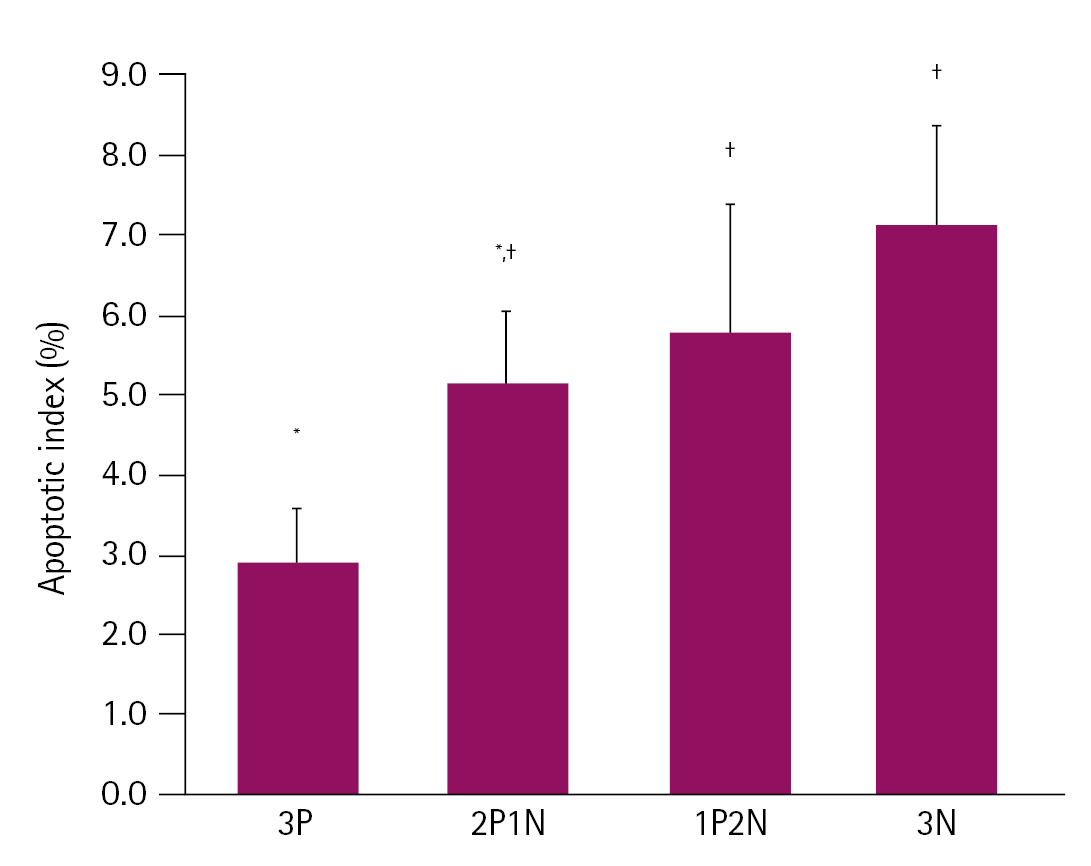
In the fourth experiment, we investigated the proportion of apoptotic cells in aggregate-derived blastocysts. When blastomeres were aggregated using the PHA-L, the apoptotic cell ratio of the blastocysts was significantly higher in the 1P2N group and 3N group compared to that in the 3P group (5.91%-6.46% vs. 2.94%, respectively). However, no significant difference was observed between the 2P1N group and other treatment groups (Fig. 3).
Discussion
Pig is an animal attracting attention as a human disease and xenotransplantation model [20]. However, the low quality of NT-derived embryos and piglet production efficiency are obstacles [21,22]. A high-quality blastocyst production technique through the aggregation of blastomeres can improve the quality of NT-derived embryos with low developmental ability. In this study, we investigated the preimplantation developmental competence and cell death of blastocysts generated through the aggregation of PA- and NT-derived oocytes.
ZP removal is an essential process for blastomere aggregation. Establishing more reliable methods for ZP removal is vital to improve embryo quality and expand its application in artificial reproduction techniques. The effect of ZP removal using TS and PE on blastomere aggregation efficiency in pigs has not been reported. Several studies have demonstrated that the presence of ZP is not required for embryonic development in vitro [23,24]. In a previous study, ZP removal using PE was successful in patients undergoing oocyte donation cycles after two or more failed attempts [25]. TE solutions are also commonly used in artificial reproduction techniques and have been used for assisted hatching and blastomere isolation in humans. In humans, ZP-free blastocysts produced using TS solutions have increased implantation rates and resulted in more successful pregnancies in patients with poor prognoses [26]. On the contrary, adverse effects of ZP removal on embryonic development have also been reported [19,27]. The method used for ZP removal has side effects such as damage to embryos due to toxicity and separation of blastomeres during division [15,27]. In our study, both the TS and PE methods showed a high survival rate of 96.0% to 97.9% after the removal of ZP. However, the ZP removal using the PE method demonstrated a significantly lower blastomere cohesion rate than the TS method (69.2% vs. 88.3%; p < 0.05). In embryos in which the ZP was removed by PE, it can be assumed that the remaining invisible ZP interfered with the cytoplasm-cytoplasm contact in our aggregation method using hemagglutinin. However, the blastocyst development rate of normally aggregated embryos developed using the two treatment methods did not differ significantly. Removal of the ZP of a two-cell stage embryo showed the side effect of separating two blastomeres during the culture process. However, the strong cell aggregation activity of PHA-L could inhibit the separation of blastomeres.
Blastomere aggregation is essential to produce chimeric animals using early-stage embryos (2-cell stage to morula) [11,28]. In several mammals, embryo aggregation techniques are utilized to increase production efficiency by correcting developmental problems within in vitro embryos. In particular, it has been observed that pigs respond well to embryo aggregation, increasing the rate at which blastocysts form, the total number of cells, ratio of inner cell mass to trophectoderm cells, and survivability of the cells [13,14]. Blastomere aggregation improves embryonic development by decreasing the level of cell death and improving cell reprogramming [29,30]. Piglets were successfully produced through the aggregation of NT embryos [31]. In our study, aggregated NT-derived blastocysts showed significantly larger diameters (181-195 μm vs. 147 μm) than control cells (non-aggregated blastomeres) regardless of the type of aggregation method. In addition, no difference was observed in blastocyst development rate and blastocyst diameter when aggregation of blastomeres was induced in NT-derived embryos using WOW and PHA-L techniques. This result is consistent with our previous study when the blastomere of PA-derived embryos was aggregated [12]. The WOW method is most frequently utilized for embryo aggregation. It has been reported that the WOW method of aggregating NT-derived blastomeres can successfully create high-quality blastocysts in pigs [30]. Nevertheless, no commercially available micro-well dish exclusively for the WOW method exists. Therefore, researchers usually make their own micro-wells for WOW and use them for experiments [32-34], and the non-uniform size of micro-wells may affect consistent blastocyst production. In addition, hand-made WOW dishes may damage embryos due to the rough surface of the micro-wells and may damage the blastocysts firmly attached to the bottom during separation [12]. PHA-L can be a suitable embryo aggregation method to replace the WOW method. In a previous study, we improved the aggregation efficiency of PA-derived embryos using PHA-L, confirmed the establishment efficiency of embryonic stem cell-like cells, and completed the differentiation capacity of embryoid bodies to three-germ layers [12].
We investigated whether support through the aggregation of PA-derived blastomeres could improve the preimplantation embryonic development of NT-derived embryos using the PHA-L method. Mammalian PA embryos invariably die in mid-gestation from imprinted gene defects and placental hypoplasia. Based on chimera experiments, trophoblastic proliferation is supposed to be inhibited in the absence of a male genome [35]. However, blastomere aggregation of IVF-derived and PA-derived embryos can produce chimeric offspring in mice and cattle [9,36]. A study by Paldi et al. [36] reported that PA-derived cells are not cell lethal and can participate to some degree in the normal differentiation of certain tissues; however, their extensive presence reduces the viability of chimeric animals and retard their postnatal development. Interestingly, our results indicate that the quality of chimeric blastocysts improved when the embryo was aggregated with a high rate of PA-derived blastomeres. In particular, the 2P1N treatment group showed a significant increase in the rate of blastocysts compared to the 3N treatment group, and the diameter of the blastocysts was considerably increased. When the apoptotic cell rate in the blastocyst was investigated, the 3P-treated group showed a significantly lower apoptotic cell rate than the 3N-treated group. In addition, it was confirmed that the cell death rate decreased when the ratio of PA-derived blastomeres in aggregates increased. GFP-transfected cells were used as NT donor nuclei to differentiate between PA-derived blastomeres and NT-derived blastomeres. Considering the possibility that PA-derived blastomeres, which have a relatively high developmental potential, form blastocysts by inhibiting NT-derived blastomeres, the distribution of GFP expression at the blastocyst stage was investigated. Interestingly, the distribution of GFP was independent of the aggregation ratio of NT-derived blastomeres. In addition, it was confirmed that the distribution of GFP within the blastocyst was not limited to a specific site and emitted light from various sites. Based on these results, the aggregation of PA-derived embryos can be used to improve the attributes of low-quality preimplantation embryos.
Although no attempt was made on piglet production, these results indicate that the NT embryo aggregated with two PA embryos improved the rate of blastocysts with increased blastocyst diameter. This method can support the early development of embryos with remarkably low developmental competence, like transgenic embryos, and provide fundamental knowledge for producing chimeric animals.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print