뇌졸중에서 클로로겐산 투여에 의한 γ-enolase 감소 완화 효과
Alleviation of γ-enolase decrease by the chlorogenic acid administration in the stroke animal model
Article information

Abstract
Stroke is a major cause of death and long-term disability. Chlorogenic acid is a phenolic compound with a potent neuroprotective effect. γ-enolase is a phosphopyruvate hydratase found in mature neurons and plays an important role in the neuronal survival. This study investigated whether chlorogenic acid regulates the expression of γ-enolase during cerebral ischemia. Middle cerebral artery occlusion (MCAO) was performed to induce cerebral ischemia. Adult male rats were used and chlorogenic acid (30 mg/kg) or phosphate buffered saline (PBS) was injected intraperitoneally 2 hours after MCAO surgery. Cerebral cortical tissues were collected 24 hours after MCAO surgery. Our proteomic approach identified the reduction of γ-enolase caused by MCAO damage and the mitigation of this reduction by chlorogenic acid treatment. Results of reverse transcription-polymerase chain reaction and Western blot analyses showed a decrease in γ-enolase expression in the PBS-treated MCAO group. However, chlorogenic acid treatment attenuated this decrease. Results of immunofluorescence staining showed the change of γ-enolase by chlorogenic acid treatment. These results demonstrated that chlorogenic acid regulates the γ-enolase expression during MCAO-induced ischemia. Therefore, we suggest that chlorogenic acid mediates the neuroprotective function by regulating the γ-enolase expression in cerebral ischemia and may be used as a therapeutic agent for brain diseases including stroke.
서론
뇌졸중은 신경세포의 손상을 유발하며, 높은 치사율을 가지는 질환이다[1]. 뇌졸중은 크게 뇌혈관이 터져서 발생하는 출혈성 뇌졸중과 뇌혈관이 막혀서 발생하는 허혈성 뇌졸중으로 구분된다[2]. 허혈성 뇌졸중은 산소공급의 부족으로 인한 활성산소종(reactive oxygen species)의 생성, adenosine triphosphate 생성 감소, 염증반응 및 세포사멸을 유도하여 세포 손상을 일으킨다[3,4]. 허혈성 뇌졸중에 의한 글루코스(glucose)의 감소는 신경세포의 성장과 분화를 촉진시키는 단백질의 활성을 억제하여 신경세포의 사멸을 유도한다[4]. 또한, 허혈에 의한 글루타메이트(glutamate)의 과도한 생성은 신경독성을 유발하여, 산화 스트레스에 의한 신경세포의 손상을 유도한다[5,6].
Chlorogenic acid는 과일, 커피, 차에 풍부하게 존재하는 천연 폴리페놀 화합물 중 하나이다[7]. 다양한 식물에서 추출된 페놀산은 유용한 기능을 가지며, 특히 chlorogenic acid는 항산화, 항균, 항돌연변이, 항염증의 효능을 가진다고 알려져 있다[8]. Chlorogenic acid는 지질다당체에 의해 유도된 염증반응을 완화시키고, β-amyloid에 의해 유도된 해마(hippocampus)의 신경세포 사멸을 억제한다[9,10]. 또한, chlorogenic acid는 혈액-뇌장벽(blood-brain barrier)을 통과하는 장점을 가져 중추신경계의 신경세포를 보호하고, 신경세포의 세포사멸을 저해한다[11]. 또한, 뇌졸중 모델에서 chlorogenic acid는 산소-포도당 결핍(oxygen-glucose deprivation)으로부터 일차뉴런을 보호하고, 염증과 산화적 스트레스를 감소시켜 신경세포의 사멸을 막는다[12]. 앞선 연구에서 우리는 중간대뇌동맥폐쇄술(middle cerebral artery occlusion, MCAO)로 유도된 허혈성 뇌졸중 모델에서 chlorogenic acid가 대뇌손상으로 유도된 신경학적 행동 장애를 개선시키고, 뇌경색과 세포사멸의 정도를 완화시킴을 보고하였다[13].
Enolase는 세포의 해당(glycolysis) 과정에 관여하는 해당효소(glycolytic enzyme)로서, 세포의 성장 및 분화를 포함한 다양한 세포 활동에 관여한다[14]. Enolase는 α, β, γ의 세가지 동종효소를 가지며, 이들은 생체 내 분포 위치가 다르다고 알려져 있다[15]. α-Enolase는 대부분의 조직에 존재하는 반면, β-enolase는 근육세포에서, γ-enolase는 성숙한 신경세포와 신경내분비세포에서 발현된다[16,17]. 특히, γ-enolase는 신경특이적(neurospecific) enolase로 명명되며, 신경세포에 영양을 공급하고 신경조직을 보호하는 특성을 가진다[18,19]. γ-Enolase는 phosphatidylinositol 3-kinase (PI3K)와 mitogen-activated protein kinase (MAPK) 신호전달경로를 활성화하여 신경세포의 성장, 분화, 재생을 조절하며, 염증성 사이토카인을 촉진시켜 신경 염증반응을 조절한다고 알려져 있다[20,21]. 또한, γ-enolase는 정상 조직에서는 세포 내에 있지만 세포 손상에 의해 세포 외 공간으로 방출되어 손상으로부터 신경세포를 보호한다[22]. Chlorogenic acid는 허혈성 뇌손상 시, 신경세포의 손상을 완화시켜 신경보호 효과를 나타내지만, chlorogenic acid의 γ-enolase 발현 조절 여부에 관한 연구는 아직까지 보고되지 않았다. 따라서, 본 연구에서는 MCAO로 유도된 허혈성 뇌손상 모델에서 chlorogenic acid 투여에 의해 변화하는 단백질 중 γ-enolase 발현의 변화를 조사하고자 하였다.
재료 및 방법
실험동물 및 실험설계
실험동물은 (주)샘타코바이오코리아(Korea)에서 구입한 Spargue-Dawley 수컷 흰쥐(7주령, 210-220 g, n = 64)로, 구입 후 일주일 동안 온도(25℃), 습도(50% ± 5%), 빛(12 hours/12 hours 명/암)이 조절되는 동물실에서 사육되었다. 본 실험은 phosphate buffered saline (PBS)을 투여한 정상군(PBS + sham군), chlorogenic acid를 투여한 정상군(chlorogenic acid + sham군), PBS를 투여한 MCAO군(PBS + MCAO군), chlorogenic acid를 투여한 MCAO군(chlorogenic acid + MCAO군)으로 나누어 진행되었다. 모든 동물실험 절차는 경상대학교 동물실험윤리위원회(Institutional Animal Care and Use Committee, IACUC)의 승인을 받아 운영 지침에 따라 수행되었다(GNU-220222-R0021).
약물 투여
용매제로서 PBS가 사용되었고, chlorogenic acid (Sigma-Aldrich, USA)는 PBS에 녹여 준비되었다. 실험동물들은 MCAO 수술 2시간 전에 PBS 또는 chlorogenic acid를 복강 안으로 주입되었다. 투여된 chlorogenic acid의 양은 30 mg/kg으로 앞서 보고된 논문을 참고하여 결정되었으며, chlorogenic acid를 투여하지 않은 동물은 PBS만을 단독 투여하였다[23]. 실험기간 동안 모든 동물들은 물과 사료가 자유롭게 공급되는 환경에서 사육되었다.
중간대뇌동맥폐쇄술
MCAO는 허혈성 대뇌손상을 유도하기 위해 실시되었고 앞서 보고된 방법에 따라 수행되었다[13]. 실험동물은 Zoletil (50 mg/kg; Virbac, France)을 근육 내로 주입하여 마취되었고, 목의 배쪽 정중선을 절개하여 오른쪽 온목동맥(common carotid artery)을 노출시켰다. 노출시킨 혈관을 주변 조직으로부터 분리한 후, 오른쪽 바깥목동맥(exteral carotid artery), 오른쪽 속목동맥(internal carotid artery)을 노출시켰다. 오른쪽 바깥목동맥의 끝부분을 6/0 봉합사로 묶고 오른쪽 온목동맥은 microvascular clip을 이용하여 혈류를 차단한 후, 오른쪽 바깥목동맥을 절제하였다. 오른쪽 바깥목동맥의 절제 부위로, 가열로 끝부분을 둥글게 만든 4/0 nylon monofilament를 삽입하여 약간의 저항이 느껴질 때까지 오른쪽 속목동맥을 향하여 밀어 넣었다. 삽입된 nylon monofilament의 길이는 약 22-25 mm였으며, filament는 오른쪽 중간대뇌동맥을 폐쇄하여 혈액의 흐름을 차단시켰다. 오른쪽 온목동맥을 차단했던 microvascular clip을 조심스럽게 제거하였다. MCAO를 실시하지 않은 sham군은 nylon filament 삽입을 제외하고는 MCAO군과 동일한 방법으로 진행되었다. 실험동물은 filament 삽입 24시간 후에 안락사되었고, 대뇌피질 조직을 채취하여 실험전까지 -70℃에서 보관하였다.
신경학적 결손 점수 검사
신경학적 행동 장애는 신경학적 결손 점수 검사(neurological deficit scoring test)에 의해 평가되었다[24]. MCAO 수술 24시간 후에 신경학적 결손 점수 검사를 실시고, 점수는 0-4점으로 측정되었다. 신경학적 결손이 없는 정상적인 자세는 0점(신경학적 결손 없음), 반대쪽 앞다리의 불완전한 연장 자세는 1점(경도의 신경학적 결손), 반대쪽으로 자발적으로 선회하는 행동은 2점(중도의 신경학적 결손), 반대쪽으로 떨어지거나 자극에 대한 민감한 반응 및 발작은 3점(심각한 신경학적 결손), 자발적 움직임이 없고 의식이 없는 상태는 4점(매우 심각한 신경학적 결손)으로 구분되었다.
코너 테스트
코너 테스트(corner test)는 앞서 보고된 방법으로 실시되었다[25]. 실험동물은 30 × 20 × 1 cm3 크기의 두 판 사이에 놓였다. 30°로 두 판의 가장자리를 연결하였고, 실험동물들을 모서리로 이동시켰다. 실험동물이 판의 구석에 도달했을 때, 양쪽 콧수염은 동시에 판에 닿았고, 동물들은 열린 끝을 향하여 뒤로 돌았다. 동물들은 MCAO 수술 전 7일 동안 훈련을 받았고, 오른쪽과 왼쪽으로 회전하는 비율이 비슷한 동물만을 본 실험에 사용했다. 코너 테스트는 실험동물 당 10회씩 측정하였으며 실험결과는 오른쪽과 왼쪽 회전 횟수로 나타내었다.
이차원겔 전기영동
오른쪽 대뇌피질 조직을 lysis buffer (8 M urea, 4% 3-[(3-Cholamidopropyl) dimethylammonium]-1-propanesulfonate [CHAPS], ampholytes and 40 mM Tris-HCl)에 넣어 균질화한 후, 원심분리기(16,000g, 4℃)로 20분간 원심분리하여, 상층액을 회수하였다. 단백질 시료는 Bradford 시약(Bio-Rad, USA)을 사용하여 소혈청알부민(bovine serum albumin)을 기준으로 정량하였으며, 이차원겔 전기영동(2-dimensional gel electrophoresis)을 이용하여 단백질 분석을 진행하였다. 먼저, first-dimensional electrophoresis를 수행하기 위해 immobilized pH gradient (IPG) gel strips (17 cm; Bio-Rad)에 단백질 시료 50 ㎍과 sample buffer (8 M urea, 2% CHAPS, 20 mM dithiothreitol (DTT), 0.5% IPG buffer, bromophenol blue)를 넣어 13시간 동안 상온에서 반응시켰다. Ettan IPGphor 3 System (GE Healthcare, Sweden)을 이용하여 strips를 250 V로 15분, 10,000 V로 3시간, 10,000 V에서 50,000 V로 상승할 때까지 focusing을 진행한 후, 1% DTT가 포함된 equilibration buffer (6 M urea, 30% glycerol, 2% sodium dodecyl sulfate (SDS), 50 mM Tris-HCl [pH 8.8])에서 15분, 2.5% iodoacetamide가 포함된 equilibration buffer에서 15분 반응시켰다. Stip은 SDS-polyacrylamide gradient gels (7.5%-17.5%)에 올려 이차원겔 전기영동이 진행되었고, bromophenol blue dye가 gel의 바닥에 도달할 때까지 Protein-Ⅱ Ⅺ electrophoresis equipment (Bio-Rad)를 이용하여 10℃에서 5 mA로 2시간, 10 mA로 10시간 전기영동되었다. Gel은 고정액(12% acetic acid and 50% methanol)에서 2시간 고정되었고, 50% ethanol에 20분간 반응시켰다. Gel은 silver solution (0.2% silver nitrate)에서 20분 염색되었고, 현상액(0.2% sodium carbonate)으로 현상 된 후 Agfar ARCUS 1200 스캐너(Agfa-Gevaert, Belgium)를 이용하여 이미지 파일로 저장되었다. 이미지는 PDQuest 2-D analysis software (Bio-Rad)로 분석되었고, 표적 단백질 spot을 떼어내어, trypsin이 함유한 완충액에 반응시킨 후 matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF)를 진행하였다. 추출한 단백질에 대하여 Voyager System DE-STR MALDI-TOF mass spectrometer (Life Technologies, Carlsbad, CA, USA)를 이용하여 mass spectrometry를 실시하였고, MS-FIT과 ProFound program을 이용하여 단백질의 정보를 추적하고, NCBI database를 이용하여 단백질 서열을 확인하였다.
Reverse transcription- polymerase chain reaction
오른쪽 대뇌피질 조직은 Trizol Reagent (Life Technologies, USA)에 넣어 균질화되었고, 원심분리기(13,000g, 4℃)를 이용하여 20분간 원심분리한 후 상측액에서 total RNA를 추출하였다. Reverse transcription-polymerase chain reaction (PCR)은 superscript III firststrand system (Invitrogen, USA)을 이용하여 수행되었고, 방법은 제조사의 지침에 따라 total DNA를 역전사시켜 단일 가닥 상보적 DNA를 얻었고, PCR을 통해 cDNA의 γ-enolase의 유전자 서열을 증폭시켰다. 사용된 γ-enolase의 primer 염기서열은 5'-TGGATCTCCATACTG CCAAAG-3' (forward)과 5'-CCAACTCCTCTTCAA TCCTCAT-3' (reverse)였다. β-actin의 primer 염기서열은 5’-GGGTCAGAAGGACTCCTACG-3’(forward)과 5’-GGTCTCAAACATGATCTGGG-3’(reverse)였다. PCR 반응은 94℃에서 5분 denature 과정, 94℃에서 30초, 54℃에서 30초, 72℃에서 1분의 30 cycles 과정을 거쳤으며, 이후 72℃에서 10분 enlongation 과정으로 시행되었다. PCR products는 Loading STAR (Dyne Bio, Korea)와 혼합하여 1% agarose gel에서 전기영동되었고, 분리된 PCR product bands를 ultraviolet-light에서 관찰한 후, SigmaGel 1.0 (Jandel Scientific, USA)와 SigmaPlot 4.0 (SPSS Inc., USA) 프로그램으로 분석되었다. γ-Enolase 발현 수준은 β-actin의 발현 강도에 대한 γ-enolase의 강도로 평가되었다.
Western blot 분석
오른쪽 대뇌피질 조직은 lysis buffer (1% Triton X-100 and 1 mM EDTA in phosphate buffer saline)에 넣어 분쇄기(homogenizer)로 분쇄되었고, 분쇄된 혼합물은 원심분리기(15,000g, 4℃)를 이용하여 20분간 원심분리한 후 상층액은 회수되었다. 단백질 측정은 bicinchoninic acid protein assay kit (Pierce, USA)를 이용하였고, 소혈청알부민을 기준으로 단백질 시료의 정량이 이루어졌다. SDS-polyacrylamide gel에 단백질 시료(30 ㎍)를 넣어 전기영동한 후, 단백질들은 gel로부터 poly-vinylidene fluoride membrane (Millipore, USA)으로 옮겨졌다. 비특이적인 항체반응을 억제하기 위하여 5% skim milk 용액에 membrane은 1시간 동안 반응되었고, tris-buffered saline with 0.1% Tween 20 (TBST)로 5분간 3회 세척되었다. 1차 항체로서 γ-enolase (1:1,000; Santa Cruz Biotechnology, USA) 또는 β-actin (1:1,000; Santa Cruz Biotechnology)을 1시간 동안 반응시켰고, TBST로 3회 세척한 후, horseradish peroxidase-conjugated goat anti-rabbit IgG (1:5,000, Pierce)를 2차 항체로 처리하였다. TBST로 세척한 후 membrane을 enhanced chemiluminescence Western blot analysis system (Amersham Pharmacia Biotech, USA)에 1분간 반응시킨 후, X-ray film에 1분간 노출시켰고, 현상액에서 현상한 후 고정액으로 고정하였다. SigmaGel 1.0과 SigmaPlot 4.0을 이용하여 발색을 확인하고 단백질 발현양을 측정하였다. γ-Enolase 발현 수준은 β-actin의 발현 강도에 대한 γ-enolase의 강도로 평가되었다.
면역형광염색
적출된 뇌조직을 4% 완충 포르말린 고정액에서 24시간 고정시켰다. 고정된 조직은 수세 후 ethanol (70%-100%)과 xylene으로 수세 및 투명화 과정을 거쳤다. 파라핀 포매기(paraffin embedding center; Leica, Germany)에서 포매한 후, 회전식 박절기(Leica)를 이용하여 4 μm의 두께로 박절하였다. 박절된 조직은 슬라이드글라스 위에 올려졌고, 신전기(slide warmer; Thermo Fisher Scientific, USA)에서 건조되었다. 박절된 조직은 xylene으로 탈파라핀되었고, ethanol (100%-70%)을 이용하여 함수화 과정을 거쳤다. 수세된 조직은 항원을 노출시키기 위해 전자레인지를 이용하여 0.1 M sodium citrate buffer (pH 6.0)에 반응시켰고, 비특이적인 항체의 결합을 억제하기 위해 0.5% fetal bovine serum에 1시간 처리되었다. 1차 항체로 anti-γ-enolase (1:100; Santa Cruz Biotechnology)을 4℃에서 12시간 반응시켰고, PBS로 3회 세척한 후, 2차 항체로서 fluorescein isothiocyanate-conjugated secondary antibody (1:100; Santa Cruz Biotechnology)를 실온에서 1시간 반응시켰다. PBS로 세척 후, 핵은 4',6-diamidino-2-phenylindole (DAPI; Santa Cruz Biotechnology)로 염색되었고, 수세 후 Ultra Cruz mounting medium을 이용하여 조직을 봉입하였다. 공초점 현미경(confocal microscope, FV- 1000; Olympus, Japan)을 이용하여 염색된 부위를 확인하였고, 결과 분석을 위하여 오른쪽 대뇌피질의 5개 부위를 무작위로 선택하였다. 대뇌피질의 이미지가 캡처되었고, Image-Pro Plus image analysis software로 상대적인 통합 밀도(relative integrated density)를 측정하였다. γ-Enolase 발현 수준은 PBS + sham군의 통합 강도에 대한 각 실험군의 통합 강도의 비율로 나타내었다.
통계분석
모든 실험결과는 평균 ± 표준편차로 표시하였으며, 실험군 간의 차이는 two-way ANOVA와 post-hoc Scheffe’s test로 분석되었다. p<0.05인 경우 통계적으로 유의한 것으로 간주하였다.
결과
MCAO 손상 시 chlorogenic acid 투여에 의한 신경행동학적 장애 개선 효과
먼저 γ-enolase 발현의 변화를 조사하기전에 MCAO 손상 시 chlorogenic acid의 신경행동학적 장애의 개선을 확인하였다. MCAO 손상은 심각한 신경학적 행동 장애를 나타내었다. PBS 투여군에서는 한 방향으로 회전, 움직임 부족, 발작 등 심각한 행동 장애를 보였으나, chlorogenic acid의 투여는 이러한 장애를 개선시켰다. Sham군에서는 PBS와 chlorogenic acid의 투여와 상관없이 행동 장애 증상이 나타나지 않았다(Table 1). 감각운동의 방향 패턴을 평가하기 위해 코너 테스트를 수행하였고, MCAO 수술군은 오른쪽 뇌손상으로 인하여 오른쪽으로만 반응을 보이는 행동 장애를 나타냈었고, chlorogenic acid의 투여는 이러한 반응을 완화시켰다(Table 1). 본 신경학적 행동 테스트는 MCAO를 통한 허혈성 뇌졸중 동물모델이 잘 확립이 되었는지를 확인하고자 신경학적 행동 검사를 실시하였고, chlorogenic acid에 의한 신경보호 효과를 확인하는 기본 자료를 제시하고자, 결과 부분에 삽입하였다.

Neurobehavioral scoring and corner tests in the in the PBS + MCAO, CGA + MCAO, PBS + sham, and chlorogenic acid + sham groups
MCAO 손상 시 chlorogenic acid 투여에 의한 γ-enolase 발현 감소의 완화
프로테오믹스 분석법을 기반으로 MCAO에 의해 유도된 대뇌허혈 시에 chlorogenic acid 투여에 의한 γ-enolase 단백질의 발현 변화를 관찰하였다. γ-Enolase의 peptide mass는 14/70이었고 sequence coverage는 34%였다. MCAO 수술은 대뇌피질의 γ-enolase 단백질 발현 감소를 유도하였고, MCAO에 의한 γ-enolase 발현의 감소는 chlorogenic acid의 투여로 완화되었다(Fig. 1A). 그러나 수술과정에서 nylon을 삽입하지 않은 sham군에서는 PBS와 chlorogenic acid 투여와 상관없이 γ-enolase의 발현은 유사하게 나타났다. γ-Enolase 발현 정도는 PBS + sham군의 결과에 대한 비율로 평가되었고, PBS + MCAO군에서는 0.48 ± 0.03이었고, chlorogenic acid + MCAO군에서는 0.93 ± 0.04였다(Fig. 1B). Reverse transcription-PCR과 Western blot 분석은 chlorogenic acid 투여가 MCAO 손상으로 유도된 γ-enolase 발현의 감소를 완화시킴을 보여주었다(Figs. 2, 3). γ-Enolase mRNA 발현은 MCAO 손상으로 감소되었고, chlorogenic acid 투여는 이들 감소를 완화시켰다(Fig. 2A). γ-Enolase mRNA 발현은 PBS 또는 chlorogenic acid을 처리한 sham군에서는 유사하게 나타났으며, PBS + MCAO군에서는 0.50 ± 0.02였고, chlorogenic acid + MCAO군에서는 0.77 ± 0.02였다(Fig. 2B). γ-Enolase 단백질의 발현은 PBS + MCAO군과 chlorogenic acid + MCAO군에서 각각 0.66 ± 0.03과 0.95 ± 0.04로 나타났다(Fig. 3A and B). 면역형광염색(immunofluorescence stain) 결과는 MCAO 손상 시에 chlorogenic acid 투여에 의한 γ-enolase 발현의 변화를 보여주었다(Fig. 4A). 대뇌피질 부위의 γ-enolase의 강도는 sham군에 비해 PBS + MCAO군에서는 유의하게 감소하였고, chlorogenic acid + MCAO군에서는 MCAO 손상으로 유도된 감소를 완화시켰다(Fig. 4A). 면역형광염색을 통한 γ-enolase 발현은 PBS + MCAO군과 chlorogenic acid + MCAO군에서 각각 0.34 ± 0.04와 0.69 ± 0.06으로 나타났다(Fig. 4B).
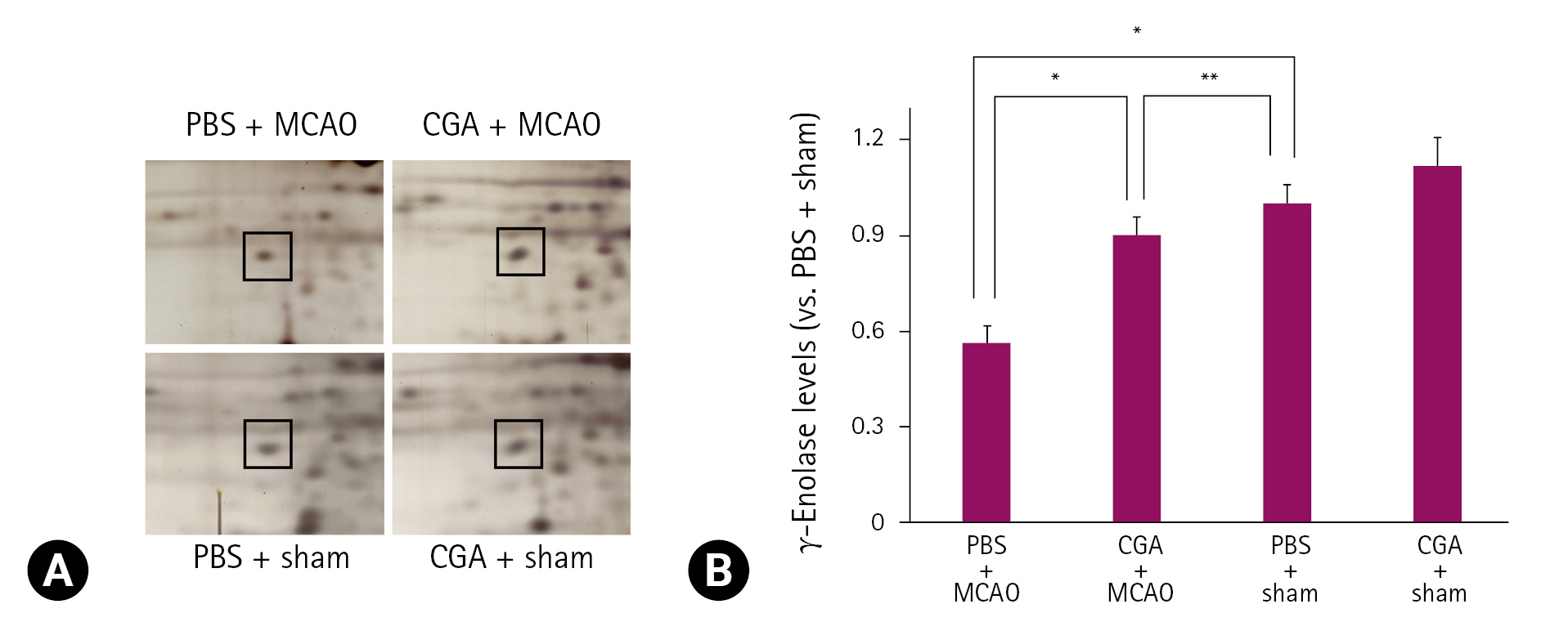
γ-Enolase protein spots identified by MALDI-TOF in the cerebral cortex of phosphate buffered saline (PBS) + middle cerebral artery occlusion (MCAO), chlorogenic acid + MCAO, PBS + sham, and CGA + sham groups. Squares indicate the γ-enolase protein spots (A). The intensity of spots was measured using PDQuest software (B). γ-Enolase level is described as spot intensity of each groups to spot intensity of PBS + sham groups. Data (n = 4) are shown as mean ± standard error of the mean. *p < 0.01, and **p < 0.05.
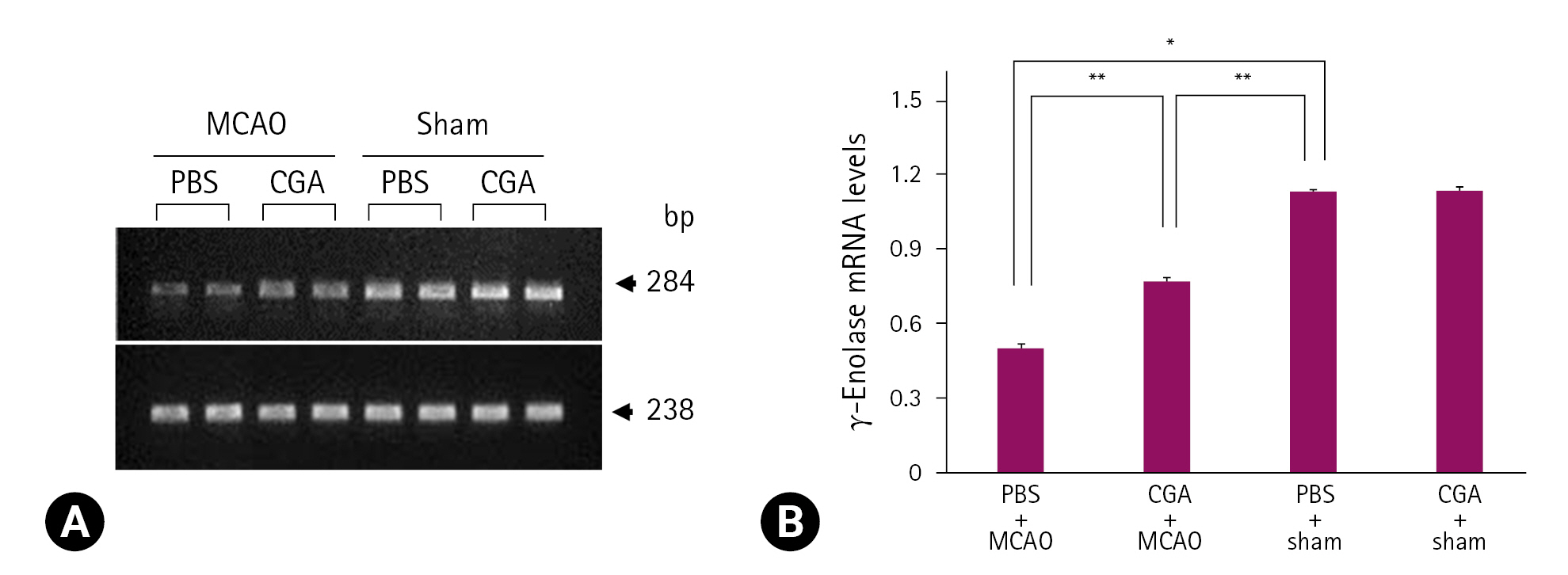
Reverse transcription-polymerase chain reaction analysis of γ-enolase mRNA levels in the cerebral cortex of phosphate buffered saline (PBS) + middle cerebral artery occlusion (MCAO), chlorogenic acid + MCAO, PBS + sham, and chlorogenic acid + sham groups. Each lane represents an individual animal (A). Densitometric analysis is represented as intensity of γ-enolase to intensity of β-actin (B). Data (n = 4) are shown as mean ± standard error of the mean. *p < 0.01, and **p < 0.05.

Western blot analysis of γ-enolase protein levels in the cerebral cortex of phosphate buffered saline (PBS) + middle cerebral artery occlusion (MCAO), chlorogenic acid + MCAO, PBS + sham, and chlorogenic acid + sham groups. Each lane represents an individual animal (A). Densitometric analysisis represented as intensity of γ-enolase to intensity of β-actin (B). Data (n = 4) are shown as mean ± standard error of the mean. *p < 0.01, and **p < 0.05.

Imunofluorescence staining with γ-enolase (green color) and DAPI (nuclei marker, blue) in the cerebral cortex of phosphate buffered saline (PBS) + middle cerebral artery occlusion (MCAO), chlorogenic acid + MCAO, PBS + sham, chlorogenic acid + sham groups (A). Arrows indicate the γ-enolase positive cells. Scale bars = 100 μm. γ-enolase relative integrated density is described as the density of each groups to the density of PBS + sham groups (B). Data (n = 4) are shown as mean ± standard error of the mean. *p < 0.01, and **p < 0.05.
고찰
본 연구에서는 신경학적 결손 점수 검사와 코너 테스트를 이용하여 MCAO가 심각한 신경학적 행동 장애를 일으킨다는 것을 확인하였고, chlorogenic acid의 투여는 MCAO 손상으로 유도된 행동 장애를 개선시켰다. MCAO 손상은 인지 기능 및 반대쪽 운동의 손상을 포함한 행동 장애와 대뇌피질의 조직병리학적 변화를 일으킨다고 알려져 있고, 앞선 연구에서 우리는 다양한 신경행동학적 테스트와 조직학적 조사를 통해 허혈성 뇌졸중 동물모델에서 chlorogenic acid의 신경보호 효과를 보고하였다[26]. Chlorogenic acid 투여는 MCAO 손상에 유도된 신경행동의 기능적 결함을 완화시켰고, 뇌경색의 부피를 감소시켰으며, 대뇌피질의 조직병리학적 변화와 세포사멸의 정도를 개선시켜, 허혈 손상으로부터 신경세포를 보호하였다[26]. 또한, chlorogenic acid의 투여는 대뇌허혈로 유도된 산화적 스트레스를 감소시켜 MCAO 손상으로부터 신경세포를 보호하였다[13]. 산화적 스트레스는 뇌손상의 주요 원인이며, 특히 신경세포는 높은 대사율과 항산화 방어체계가 미약하여 산화적 스트레스에 매우 취약하다고 알려져 있다[27,28]. 우리는 뇌졸중 모델에서 chlorogenic acid의 투여가 미세아교세포의 활성화를 억제하고, pro-inflammtory 단백질인 nuclear factor kappa B, interleukin-1β, tumor necrosis factor α를 억제하여 항염증효과를 나타냄을 보여주었다[14]. 또한, 대뇌허혈 손상 시 chlorogenic acid의 신경보호 작용은 다양한 단백질의 조절과정을 통하여 이루어질 것으로 추측하여, chlorogenic acid에 의해 변화되는 단백질들을 프로테오믹스 분석법으로 탐색하였다. 탐색된 단백질 중 본 연구에서는 신경세포의 다양한 세포 활동에 관여하는 γ-enolase 발현의 변화에 중점을 두었다. MCAO 손상은 γ-enolase의 발현을 감소시켰고, chlorogenic acid 투여는 이러한 감소를 완화시킨다는 것을 본 연구를 통해 확인하였다.
γ-Enolase는 신경돌기의 재생과 신경세포의 생존에 관여하며, 신경보호 효과를 나타내었다[18,19]. 또한, γ-enolase는 PI3K/Akt와 MAPK/ERK 신호전달경로의 활성화하여 신경세포의 유지에 관여하는 것으로 알려져 있어, 손상으로부터 신경세포를 보호하는 데 중요한 역할을 한다[29]. 과발현된 γ-enolase는 신경세포의 성장을 촉진하고 축삭돌기의 재생에 관여하여 신경세포의 보호제로서 작용하였다[19,30]. 따라서, 해당과정에서 에너지 생성에 중요한 역할을 하는 γ-enolase의 감소는 신경세포의 에너지 대사과정을 방해한다고 볼 수 있다. 우리는 앞서 대뇌허혈과 글루타메이트 투여로 유도된 신생아의 대뇌피질 손상 시 γ-enolase의 발현 감소를 보고하였다[31,32]. 또한, 배양된 해마신경세포에서 글루타메이트 처리는 신경세포의 사멸을 유도했고, γ-enolase의 발현을 현저히 감소시켰다. 이 연구들은 허혈과 독성물질로 인한 γ-enolase의 발현 감소가 신경세포의 손상을 유도함을 나타내었다. 본 연구에서는 프로테오믹스 분석법으로 chlorogenic acid 투여에 의한 γ-enolase의 변화를 일차적으로 확인하였고, reverse transverse-PCR과 Western blot 분석을 통해 명확하게 확인하였다. MCAO 손상으로 유도된 대뇌허혈 모델에서 γ-enolase의 발현 감소와 chlorogenic acid의 투여에 의한 γ-enolase 발현의 감소 완화를 확인하였다. 최근, 우리는 chlorogenic acid가 대뇌허혈 손상으로 유도된 인산화된 Akt와 인산화된 Bad의 발현 수준의 감소를 완화시켰으며, 인산화된 Bad와 14-3-3 단백질의 결합력의 감소를 개선시켰고, 세포사멸의 기전을 억제함을 보고하였다[33]. 이들 결과는 chlorogenic acid가 PI3K/Akt 신호전달체계를 활성화하여 대뇌허혈로 인한 손상 시 신경세포를 보호함을 나타낸다. γ-enolase는 PI3K/Akt의 신호전달경로의 활성화로 신경세포의 재생 및 신경세포의 보호작용에 관여한다고 알려져있다[30]. 이들 보고에 따르면, chlorogenic acid가 γ-enolase를 조절하고 PI3K/Akt의 신호전달경로를 활성화하여 세포의 사멸을 억제함으로써 신경세포의 생존 및 보호 기능에 관여한다고 볼 수 있다. 따라서 chlorogenic acid은 MCAO에 의해 유도되는 허혈성 손상에 대해 γ-enolase 발현을 유지하며, 이러한 γ-enolase 발현의 유지는 신경세포의 보호작용에 관여한다. Chlorogenic acid는 항산화, 항암, 항염증 등 다양한 약리학적 특성을 가지고 있으며, 임상적으로 고혈압 환자의 혈압을 감소시키는 효과가 있다고 알려져 있다[34]. 또한, chlorogenic acid는 독성으로 인한 심각한 부작용을 일으키지 않기 때문에 안전하고 효과적인 물질로 보고되어, 임상적 응용을 위해 중요한 물질로 간주되고 있다[35]. 이들 연구들을 토대로 chlorogenic acid는 뇌허혈을 포함한 염증성 신경질환의 치료에도 적용 가능한 물질로 판단된다. Chlorogenic acid에 의한 γ-enolase의 조절 기전은 아직 보고되어 있지 않지만, MCAO 손상 시 chlorogenic acid 투여에 의한 γ-enolase 발현 감소의 완화가 신경보호 효과에 관여함으로, 본 연구의 결과는 chlorogenic acid의 뇌질환 치료제로서의 활용 가능성을 제시할 수 있다. 결론적으로, 본 연구는 chlorogenic acid가 MCAO로 유도된 대뇌허혈 시 γ-enolase의 발현을 조절하여 신경세포의 보호에 관여함을 규명하였다.
Notes
The authors declare no conflict of interest.
Acknowledgements
This research was supported by the National Research Foundation of Korea (NRF) Grant funded by the Korea government (MEST) (NRF-2021R1F1A105878711).