Introduction
Feline panleukopenia virus (FPV), a member of the genus Protoparvovirus in the family Parvoviridae, is a small, non-enveloped, single-stranded DNA virus. The FPV genome contains approximately 5,200 nucleotides with 4 open reading frames (ORFs) encoding 2 non-structural proteins (NS1, NS2) and 2 structural proteins (VP1, VP2) [1]. The first ORF (which encodes the NS1 20.7 kDa polyprotein) is associated with virus replication, transcription, and host cell apoptosis [2,3]. The second ORF (which encodes NS2 of 87 amino acids) controls capsid protein assembly and translation [4]. The third ORF (which encodes VP1) contains the full sequence of VP2 and a 143-residue N-terminal sequence and is involved in the nuclear transport of capsids and efficient cell infection [5]. The 4th ORF encodes VP2; this is the major capsid protein (constituting approximately 90% of the entire capsid). VP2 controls the host range and contains several important B cell epitopes that induce protective (neutralizing) antibodies during infection [6]. Of the 4 ORFs, the one encoding VP2 has been most commonly explored in terms of genetic and epidemiological characteristics. The VP2 protein has served as the antigen of subunit vaccines [6,7].
FPV is transmitted oronasally and fecally. FPV strains have been reported in the Felidae and (globally) in wild animals, including raccoons, monkeys, and tigers [8,9]. The clinical signs of FPV infection in cats include a high fever, anorexia, vomiting, and hemorrhagic diarrhea; about half of all infected animals die [10]. Virus neutralization, hemagglutination inhibition, and enzyme-linked immunosorbent assays of canine parvovirus (CPV) have been used to detect anti-FPV antibodies in cat sera [11,12]. The methods of diagnosing FPV infection include isolation of the virus, immunofluorescence, a one-step immunochromatographic assay, enzyme-linked immunosorbent assay, and polymerase chain reaction (PCR) [13]. Although FPV infections were identified in South Korean cats in 2011 [14], isolation of the virus and genetic analyses are lacking.
In this study, we isolated 8 FPV isolates from naturally infected cats via 5 cycles of viral propagation in Crandell-Rees feline kidney (CRFK) cells. Here we define the biological features of the isolates and sequence their VP2 genes.
Materials and Methods
Samples
In total, 60 samples (5 lungs, 6 spleens, 5 brains, 4 kidneys, 4 livers, 3 hearts, 10 intestines, and 23 fecal samples) from 22 cats sent to the Animal and Plant Quarantine Agency of South Korea were used. Viruses were isolated from samples between March 2017 and October 2019. A 10% (w/v) homogenate of each sample (in Dulbecco’s modified Eagle’s medium [DMEM]) was centrifuged to remove tissue debris. All samples were passed through a 0.45-µm-pore syringe to remove bacteria.
Virus isolation, cell tropism, and titration
Samples (200 µL) were added to actively growing CRFK cells (the ATCC CCL-94 line) in 24-well plates, followed by incubation at 37°C for 1 hour. The media were removed, and 1 mL amounts of fresh DMEM with 5% (v/v) fetal bovine serum was added to each well. The plates were incubated under 5% (v/v) CO2 at 37°C for 5 days. The supernatants of cells exhibiting cytopathic effects (CPEs) were harvested and added to fresh CRFK cells (the second passage). If no CPE was observed after 5 days, samples were considered negative for the virus. To explore cell tropism, we added one (FPV19D01) of the 8 FPV isolates containing 103.0 TCID50/mL to 6 cell types growing in 25 cm2 flasks: CRFK, Fcwf-4 (ATCC, CRL2728), BHK-21 (ATCC, CCL-10), A72 (ATCC, CRL-1542), Madin-Darby canine kidney (MDCK; ATCC, CCL-34), and Vero (ATCC, CCL81) cells. After incubating for 5 days, each flask was frozen and thawed 3 times. Viral titers were determined with an indirect fluorescence assay based on the Spearman-Karber method and the results expressed as TCID50/mL.
Identification of FPV isolates via immunofluorescence assay (IFA)
CRFK cells infected with the 8 isolates containing 103.0 TCID50/mL in 96-well plates were fixed in cold 80% (v/v) acetone for 15 minutes, washed 3 times with phosphate-buffered saline (PBS), exposed to a mouse anti-CPV antibody (Median Diagnostics, Korea) at 37°C for 1 hour, and then exposed to a fluorescein isothiocyanate-conjugated goat anti-mouse immunoglobulin (Ig) G + IgM antibody (KPL Laboratories, USA). After being washed with PBS, CRFK cells were examined under a fluorescence microscope; cells evidencing intranuclear fluorescence were considered infected with FPV.
Hemagglutination assay
The hemagglutination assay (HA) assay was described previously [16]. Briefly, FPV isolates were serially diluted (in 2-fold steps) in 50 µL amounts of Sorensen buffer (pH 6.0) in a 96-well U-form microplate. Then 50 µL amounts of 0.6% (w/v) pig erythrocytes with 0.1% (w/v) bovine serum albumin in Sorensen buffer were added, followed by incubation at 4°C for 1 hour. The HA titer was the reciprocal of the highest FPV isolate dilution exhibiting an HA reaction.
Electron microscopy
Details of the electron microscopy (EM) were described previously [16]. Briefly, 2 FPV isolates (FPV19D01 and FPV19D02) propagated in CRFK cells were concentrated with polyethylene glycol (PEG) 8000 (Sigma-Aldrich, USA). The PEG-precipitated pellets were dissolved in TNE buffer (100 mM Tris-HCl, 100 mM NaCl, and 1 mM EDTA; pH 7.6) at 5% of the original volume. Two samples were placed atop 1.25 mg/mL cesium chloride (CsCl) layers in centrifuge tubes and centrifuged at 100,000 × g for 2 hours. The clear bands evident between the 1.25 and 1.45 mg/mL CsCl layers were transferred to dialysis cassettes and dialyzed overnight against PBS to remove residual CsCl. The dialyzed samples were placed into Vivaspin tubes (Sartorius, Germany) and centrifuged at 3,500 × g for 15 minutes; the supernatants were collected [17].
PCR, sequencing, and phylogenetic analyses
We extracted genomic DNA from the 8 FPV isolates using a DNA extraction kit (Bioneer, Korea) according to the manufacturer’s instructions. The DNA was subjected to PCR in terms of both diagnosis and cloning. Table 1 lists the primer sets used to diagnose FPV and the 3 sets used to amplify portions of the VP2 gene. The reaction mixture in the One-Step PCR kit (Bioneer) consisted of 5 µL denatured DNA, 1 µL of each primer (50 pmol/µL), and 43 µL distilled water. A thermal cycler (Analytik Jena, Germany) was used to perform 40 cycles of denaturation, annealing, and extension (95°C for 30 seconds, 52°C for 30 seconds, and 72°C for 30 seconds, respectively), with a final 10 minutes extension at 72°C. Approximately 10 µL of each PCR product was loaded onto 2.0% (w/v) agarose gel and subjected to electrophoresis for 30 minutes. Three PCR products (each representing a portion of the VP2 gene) were purified with a gel extraction kit and ligated into pGEM-T Easy Vector (Promega, USA) in accordance with the manufacturer’s instructions. PCR product insertions in plasmids were verified via digestion with EcoRI (Bioneer). Two plasmids per gene were sent to Macrogen (Korea) for sequencing. Three partial VP2 nucleotide sequences were determined and ligated to form the full VP2 gene. Details of the phylogenetic tree and the alignment were described previously [17].
Results
Isolation and biological characterization of FPV isolates
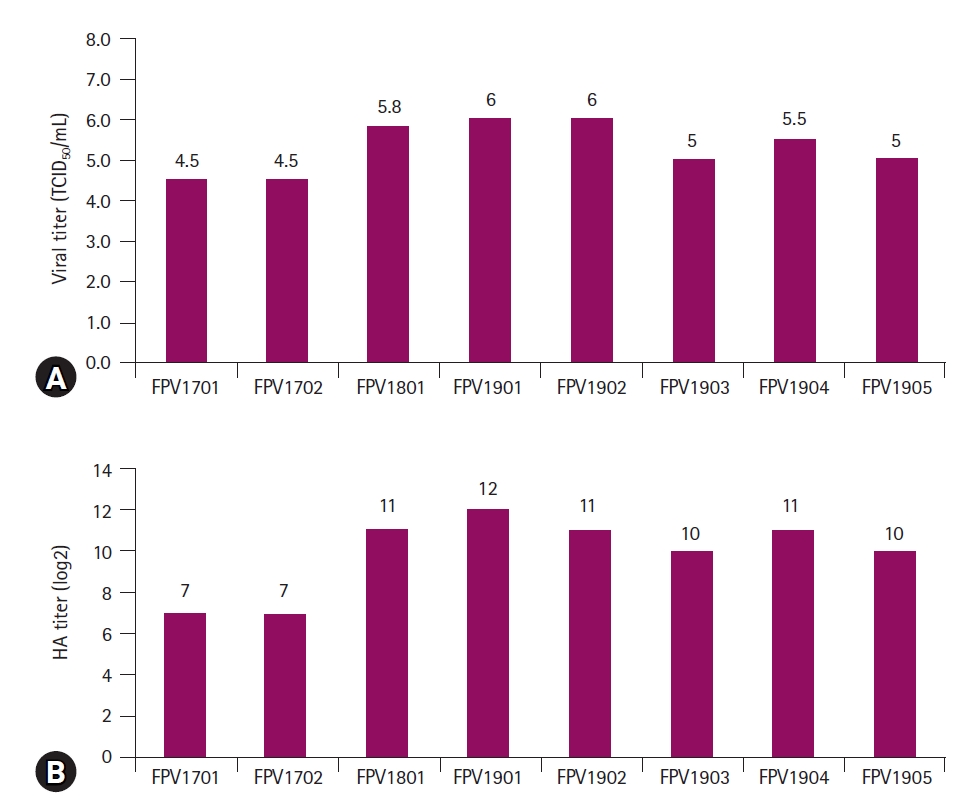
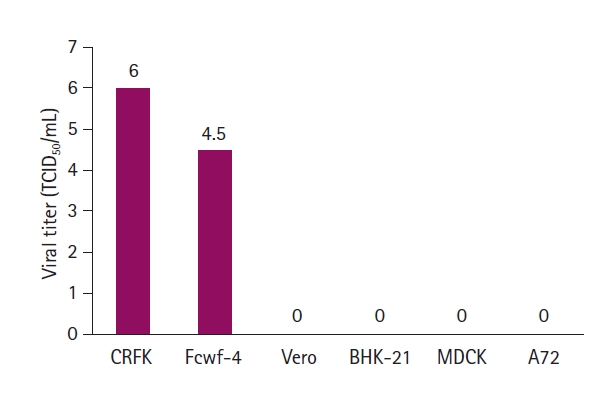
Of the 60 samples, 8 generated CPEs are characterized by the destruction of infected cells (Fig. 1A-H). The 8 virus isolates were designated FPV17D01, FPV17D02, FPV18D01, FPV19D01, FPV19D02, FPV19D03, FPV19D04, and FPV19D05. CRFK cells infected with the 8 FPV isolates were fixed in cold acetone and stained with mouse monoclonal antibodies against CPV. Fluorescence was observed in the nuclei of CRFK cells (Fig. 1I-P), which confirmed that the 8 isolates were FPVs. The growth features of all isolates were evaluated. Of the 8 isolates, FPV19D01 and FPV19D02 exhibited the highest viral titers of 106.0 TCID50/mL (Fig. 2A). The HA titers of the 8 isolates (determined using pig blood cells) ranged from 27 to 212 (Fig. 2B). All FPV isolates were inoculated into CRFK, Fcwf-4, BHK-21, A72, Vero, and MDCK cells. CRFK and Fcwf-4 cells infected with FPV developed viral titers of 106.0 and 104.5 TCID50/mL, respectively. By contrast, BHK-21, A72, Vero, and MDCK cells did not exhibit intranuclear fluorescence, which indicated an absence of viral replication (Fig. 3). Therefore, cat-derived cells were optimal for isolating FPV. Based on the FPV growth patterns, 2 isolates (FPV19D01 and FPV19D02) were propagated and harvested 5 days post-inoculation (PI), followed by EM. FPV19D01 and FPV19D02 viral particles from infected CRFK cells purified via CsCl density gradient ultracentrifugation were spherical and 20 to 30 nm in diameter and thus were typical Parvoviridae (Fig. 4).
Genetic characterization of the 8 FPV isolates
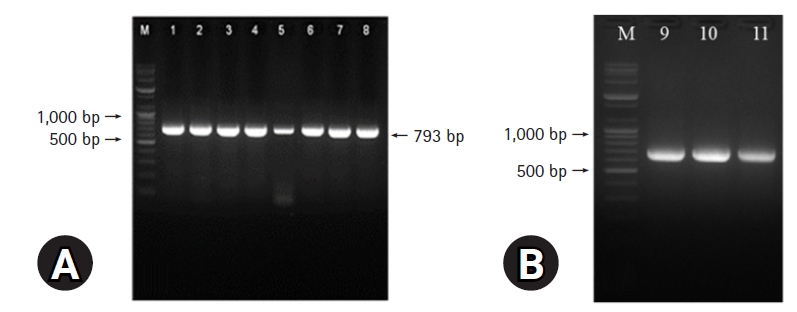
DNA extracted from the 8 FPV isolates was subjected to PCR. All PCR products were 793 bp in length, which confirmed that they originated from FPV (Fig. 5A). Three PCR products from each of the 8 FPV isolates were cloned to determine the sequences of the entire VP2 genes. Restriction enzyme digestion was used to confirm successful cloning. As shown in Fig. 5B, the 3 PCR products from FPV19D01 were 708, 719, and 736 bp in size on 2.0% (w/v) agarose gels, which verified that the isolate was indeed an FPV. The entire VP2 gene of FPV features 1,755 nucleotides encoding 585 amino acids. Comparative analyses of the complete VP2 genes of the 8 isolates revealed sequence homologies of 99.5% to 99.9% (Table 2). The VP2 genes were compared to those of 26 FPVs and 8 CPVs logged in GenBank; we explored the genetic relationships of our isolates and those of other FPVs. As shown in Fig. 6, our 8 FPV isolates were closely related to the KS42 strain (99.9% homology) isolated in Korea in 2008. Phylogenetic analyses revealed that our 8 FPV isolates fell into FPV group 3. The VP2 amino acid sequences from the 8 FPV isolates and the CPV strains were aligned with sequences from 5 FPV strains (K2, K3, K4, K50, and KS42). As shown in Table 3, the FPV18D01 and FPV19D05 isolates featured different amino acids (e.g., asparagine and isoleucine at positions 465 and 582 of the VP2 gene, respectively). The VP2 amino acid differences at position 103 (Val → Ala) and 564 (Asn → Ser) made it possible to discriminate FPV and CPV strains.
Discussion
FPV is a highly contagious viral pathogen that causes leukopenia and severe hemorrhagic diarrhea in cats [14]. FPV infections have been principally detected in the Felidae. However, such infections have rarely been reported in other animals, such as raccoons, foxes, cheetahs, wild leopard cats, or badgers [15,16]. Although cats of all ages are susceptible to FPV, young cats that lack maternal antibodies or that have not been vaccinated exhibit mortality rates of at least 50%. Although clinical symptoms (vomiting and bloody diarrhea) and one-step immunochromatography assays have greatly aided in the diagnosis of FPV infections, few reports have isolated of FPVs from Korean cats [8]. Here we isolated FPVs from 60 samples from 22 dead cats. All isolates were FPVs based on CPEs and the results of immunofluorescence assay, EM, and PCR assays.
Two feline cell lines (CRFK and Fcwf-4) were susceptible to FPV; non-feline lines (BHK-21, A72, Vero, and MDCK) were not. FPV thus preferentially replicates in feline cells. CRFK cells were optimal for isolating FPV, which suggests that FPV uses the feline transferrin receptor to enter cells [18,19]. The feline cell tropism of FPV limits its range to cats. We evaluated the biological characteristics of 8 FPV isolates; the viral titers reached 104.5 to 106.0 TCID50/mL 5 days PI, and the HA titers ranged from 27 to 212. All FPV isolates thus evidenced relatively low viral and HA titers; this may reflect biological adaptation to CRFK cells. EM of 2 FPV isolates revealed a morphology typical of FPV: FPV19D01 and FPV19D02 viral particles were of similar size and shape to parvoviruses isolated from a dog in Brazil [20].
Real-time and multiplex PCR have been used to detect FPV and differentiate it from CPV [21]. PCR amplification of partial VP2 genes from FPV isolates yielded bands of the expected size (793 bp), which confirms that all 8 isolates were FPVs. Although CPV evolution (driven by positive selection) has led to the emergence of new antigenic variants (such as CPV-2, CPV-2a, CPV-2b, and CPV-2c), FPV has evolved principally via random genetic drift and maintains its host specificity [22]. In the time since the identification of FPV in Korean cats, systematic analyses of FPV VP2 gene evolution have been lacking. We phylogenetically analysed recent FPV isolates in terms of mutation status. Phylogenetic analyses of the FPV VP2 gene revealed the evolutionary relationships and genetic variation among the FPVs [8,14,15]. Aligning the full VP2 nucleotide sequences from the 8 FPV isolates revealed that they were 99.5% to 99.9% similar. Phylogenetic analyses revealed 3 clusters of FPV strains clearly distinct from the CPV-2 strains; our 8 FPV isolates belonged to cluster G3. Six of the isolates were closely related to the KS24 strain isolated in South Korea in 2008. The high-level similarity in the VP2 gene among South Korean FPV isolates suggests that FPV does not vary temporally or geographically.
Key amino acid substitutions in VP2 allow CPV to be classified into several genotypes (termed CPV-2a/b/c). The genotypes of CPV-2 variants are generally based on different amino acids at positions 297, 300, 426, and 555 of VP2 [23]. The residue changes in VP2 that rendered it different from the FPV of CPV-2 occurred at positions 80, 90, 93, 103, 323, 564, and 568 [8]. Note that we found amino acid substitutions in the VP2 of FPV18D01 and FPV19D05 at positions 465 and 582, which indicates that a comparison of the full genomic sequences of the 2 FPV isolates is required, because such changes may affect FPV antigenicity. Although FPV strains cannot be genotyped via analyses of VP2, the differences in amino acids 103 and 564 distinguish FPV from CPV.
In conclusion, we isolated and confirmed 8 FPVs from naturally infected cats using HA, FA, EM, and PCR. CRFK cells were optimal for isolating FPV. The 8 isolates exhibited more than 99.5% VP2 gene homology. The FPV VP2 amino acid sequence data will aid in future research. More Korean FPV isolates from naturally infected cats will improve diagnosis and allow for the creation of an FPV vaccine strain. Further studies on the virulence of FPV isolates in cats are required.



 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print