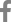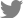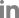Introduction
The Reoviridae family consists of the subfamilies Sedoreovirinae and Spinareovirinae. The genus Orthoreovirus in Spinareovirinae comprises five species: avian orthoreovirus, baboon orthoreovirus, mammalian reovirus (MRV), Nelson Bay orthoreovirus, and reptilian orthoreovirus [1-3]. MRVs possess 10 dsRNA segments: three large (L1-L3), three medium (M1-M3), and four small (S1-S4) segments [1]. There are four known MRV serotypes. Serotypes 1 (Lang), 2 (Jones), and 3 (Dearing) are categorized based on viral infectivity and hemagglutination activity (HA), whereas serotype 4 (Ndell) was identified using antigenic relatedness [4]. Viral neutralization and HA are associated with segment 1 (S1), which encodes the Žā1 and Žā1s proteins. The Žā1 protein is responsible for viral attachment to cell receptors, neutralization, and HA [5,6]. Therefore, molecular analysis of the S1 segment has been used to classify MRV serotypes [7,8].
The diagnosis of MRV infection is based on virus isolation, reverse transcription-polymerase chain reaction (RT-PCR), and electropherotyping via polyacrylamide gel electrophoresis [7,9-11]. MRV isolation is confirmed by its cytopathic effects (CPEs) and immunostaining with a specific MRV antibody. The typical MRV form on electropherotyping has a 3-3-4 migration pattern [9]. Sero-surveillance of MRV is conducted using virus neutralization assays and enzyme-linked immunosorbent assays [9,12].
MRVs have a broad host spectrum, and many isolates have been reported from humans and animals worldwide. Although MRVs have long been considered non-pathogenic, there is evidence that MRV infection in humans causes acute respiratory symptoms [13]. MRVs have been also associated with respiratory or enteric symptoms in animals, including acute respiratory disease in cattle [14]. Many samples of MRV serotype 3 (MRV-3) have been isolated from dogs, pigs, Chinese tree shrews (Tupaia belangeri chinensis), and civets (Paguma larvata) [7,9,14-18]. Experimental infection of neonatal pigs with MRV-3 led to acute gastroenteritis and severe diarrhea within 72 hours of infection with 100% mortality [18]. In Korea, 10 MRVs isolated from 237 pig diarrheic fecal samples were reported to belong to MRV-3 porcine lineage IV [10]. Although MRV-3 has been isolated from Korean pigs [10], there have been no reports on the isolation of Korean MRVs from wild animals.
Wild animal-derived viruses such as coronavirus disease 2019 and the highly pathogenic avian influenza virus infect humans and can cause death. In this study, a new MRV-3 isolate, designated MReo2045, was isolated from nasal swabs of a dead Korean roe deer (Capreolus pygargus) in 2020. Its biological and molecular characteristics are reported.
Materials and Methods
Samples and preparation
In this study, nasal swabs were obtained from 81 water deer (Hydropotes inermis), 43 raccoon dogs (Nyctereutes procyonoides koreensis), 16 roe deer, and four badgers (Meles meles) that had been sent to the Gangwon Wild Animal Rescue Center for treatment in 2020. Each swab was dipped in 3 mL of phosphate-buffered saline (PBS; pH 7.2). The samples were centrifuged at 2,500 ├Ś g for 15 minutes to remove tissue debris. The supernatants were filtered through a 0.45-╬╝m syringe filter and used for virus isolation.
Virus isolation, growth kinetics, and titration
Vero (CCL-81; ATCC, USA) cells used for virus isolation were cultured in DulbeccoŌĆÖs modified Eagle medium (DMEM) supplemented with an antibiotic-antimycotic (Gibco, USA) and 10% fetal bovine serum (FBS). After washing the confluent Vero cells twice with PBS, they were incubated with 100 ╬╝L of each filtered sample at 37┬░C for 1 hour. After removing the liquid in the 24-well plates, 1 mL of new DMEM containing 3% FBS was added to each well. The cells were then incubated in a 5% CO2 incubator at 37┬░C for 7 days. If the Vero cells in the second passage did not exhibit any CPEs after 7 days, the samples were considered negative for virus isolation. The supernatant in wells of cells exhibiting CPEs was harvested and inoculated into newly prepared Vero cells in a six-well plate for the second passage. The growth kinetics of the MReo2045 isolate were examined three times to determine the time of fastest growth. Briefly, Vero cells grown in 25-cm2 flasks were inoculated with an isolate at 103 50% tissue culture infectious dose (TCID50)/mL and harvested every day for 7 days. After three freeze-thaw processes, the viral titers of the isolate were determined based on the presence of typical CPEs by aliquoting 100 ╬╝L of each virus diluted 10-fold in 96-well microplates and adding 100 ╬╝L of fresh medium containing 2 ├Ś 105 Vero cells and 10% FBS to the wells. The Vero cells were observed for typical CPEs for 7 days post-inoculation. Viral titers as measured using the Spearman-Karber method were expressed in TCID50/mL.
Immunofluorescence assay
Vero cells infected with the isolate in 96-well microplates were fixed with cold 80% acetone at -20┬░C for 20 minutes. The fixed cells were reacted with a mouse anti-MRV-3 antibody at 37┬░C for 1 hour and stained with fluorescent isothiocyanate-conjugated goat-anti-mouse IgG + IgM antibodies (KPL Laboratories, USA) diluted 200-fold in PBS. Subsequently, the cells were washed three times with PBS and then examined at 200├Ś under a fluorescence microscope. Specific cytopathic fluorescence of Vero cells with CPEs was considered to indicate MRV infection.
Electron microscopy
The isolate propagated in Vero cells was concentrated and purified using a previously reported method [17]. The purified sample was subjected to negative staining with 1% uranyl acetate. Virus particles on the grid were examined under a Hitachi 7100 electron microscope (Hitachi, Japan).
HA assay
After propagation of the isolate, for the HA assay, the isolate was serially diluted two-fold with 50 ╬╝L of PBS (pH 7.2); 50 ╬╝L of 0.6% erythrocytes obtained from mice (Mus musculus), pigs, chickens, geese, and guinea pigs were then added and incubated at 4┬░C for 1 hour. The HA titer of the isolate was expressed as the reciprocal of the highest dilution of MRV that produced an HA reaction.
RT-PCR, sequencing, and phylogenetic analyses
Total genomic RNA was extracted from the isolate using an RNA extraction kit (Bioneer, Korea), in accordance with the manufacturerŌĆÖs instructions. RNA of the isolate was eluted with 50 ╬╝L of elution buffer provided in the kit. RT-PCR was performed to diagnose MRV. Table 1 lists the primers used to amplify the partial MRV S1 and S4 genes. The reaction mixture (Bioneer) comprised 5 ╬╝L of denatured RNA, 1 ╬╝L of each primer (50 pmol), and 43 ╬╝L of distilled water. The RT-PCR cycling profile consisted of complementary DNA synthesis at 50┬░C for 30 minutes; 35 cycles of 95┬░C for 30 seconds, 50┬░C for 30 seconds, and 72┬░C for 30 seconds; and a final 5-minute extension at 72┬░C. The RT-PCR products were electrophoresed on 2.0% agarose gels containing nucleic acid staining solution for 30 min and visualized using a Gel Doc EZ imager (Bio-Rad, USA).
RT-PCR products of the partial S1 and S4 genes were purified for cloning via gel extraction; each product was ligated into the pGEM┬«-T Easy vector system (Promega, USA) in accordance with the manufacturerŌĆÖs instructions. Selected white colonies grew in Luria-Bertani broth containing 50 ╬╝g/╬╝L ampicillin. Plasmid DNA was extracted from the white Escherichia coli (DH5╬▒) colonies; each DNA insert in a plasmid was identified via EcoR1 digestion. The genes were sequenced by Macrogen (Korea). The DNA sequence of each strand was cross-validated using the universal primers SP6 and T7 to verify the nucleotide sequences. The partial S1 and S4 nucleotide sequences of the MReo2045 isolate were analyzed with those of other MRVs obtained from GenBank. Phylogenetic trees based on the partial S1 and S4 genes were constructed using MEGA ver. 7.0.20 (http://megasoftware.net/). We used the bootstrap method to assess the reliability of the phylogenetic trees with 1,000 replicates.
Results
Isolation and biological characterization of the isolate
Of the Vero cells inoculated with the 144 samples, only one sample from a roe deer resulted in specific CPEs, which appeared as the cell surface peeling off and detaching (Fig. 1A). The sample with specific CPEs was named MReo2045, because it was isolated from the 45th sample collected in 2020. Vero cells infected with the isolate were fixed in cold acetone and reacted with mouse monoclonal antibodies against MRV-3. As shown in Fig. 1C, specific fluorescence was observed in the cytoplasm of Vero cells infected with MReo2045. The growth kinetics were examined to determine when the isolate had the highest titer. As shown in Fig. 2, the highest viral titer of 105.7 TCID50/mL was seen when the virus was harvested at 5 days after inoculation. HA assays were performed with five erythrocyte types (Table 2). The isolate was able to agglutinate only pig erythrocytes. Based on the MReo2045 growth kinetics, a virus antigen for electron microscopy (EM) was prepared via sucrose purification. Virus particles of the MReo2045 isolate were identified via EM from purified solution after sucrose density gradient ultracentrifugation using a sample concentrated with PEG 8000 (Sigma-Adrich, USA). The viral particles were 70 to 80 nm in size and had the typical Reoviridae morphology (Fig. 3).
Molecular characterization of the MRV isolate
PCR, cloning, and sequencing were used to determine the genetic features of the isolate. As shown in Fig. 4, 570- and 900-bp RT-PCR products from the MReo2045 isolate were detected on 2.0% agarose gels, confirming that the isolate was MRV based on the expected sizes of two RT-PCR products. The partial S1 and S4 sequences of the MReo2045 isolate comprise 900 and 570 nucleotides, respectively, encoding 300 and 170 amino acids. The partial S1 and S4 segments were compared with those of 20 MRVs listed in GenBank to determine their relationships with the other MRVs. S1 of the MReo2045 isolate had the highest homology (96.1%) with the T3/Bat/Germany/342/08 strain; S4 shared 95.9% nucleotide identity with the Slovenian strain SI-MRV06, which was isolated from bat guano. S1 of the isolate had 79.6% nucleotide identity with the KPR G strain, which was isolated from a Korean pig. Phylogenetic trees were constructed using the partial S1 and S4 segments of 20 MRVs. As shown in Fig. 5, the Korean isolate belonged to serotype 3 when the tree was constructed with S1 and was highly correlated with the SI-MRV06 and BatMRV1-IT2011 strains in the tree constructed with partial S4 gene sequences. The partial amino-acid sequences of S1 of the MReo2045 isolate, KPR G strain, and T3/Bat/Germany/342/08 strain were aligned to characterize the genetic features of the isolate. As shown in Fig. 6, three potential N-linked glycosylation sites (N-X-S/T) were identified in the partial S1 protein of the MReo2045 isolate, and 10 and 37 amino acids of the isolate were replaced in the partial S1 protein of the T3/Bat/Germany/342/08 and KPR G strains, respectively.
Discussion
MRV has a global distribution, with infections reported in humans and animals, including dogs, pigs, Chinese tree shrews, and civets [7,9,12-20]. Wild animals infected with MRV may be asymptomatic or die. Compared with other wild animal diseases, it is difficult to diagnose MRV based only on the symptoms. There are few reports on the prevalence of MRV in Korean wild animals. We collected 144 nasal swabs from wild animals sent to the Gangwon Wild Animal Rescue Center for treatment in 2020. Only one virus sample was isolated, from a nasal swab from a roe deer. When rescued, the roe deer was weak, dehydrated, and had minor injuries. During treatment, the roe deer died. We confirmed that the isolate, MReo2045, was MRV based on specific cell CPEs, immunofluorescence assay, EM, RT-PCR, and nucleotide sequence analysis.
The isolate reached 105.7 TCID50/mL at 5 days after inoculation, indicating that the MReo2045 isolate had a relatively low viral titer and was not adapted in any cell line and circulating in wild animals. The virus particles identified via EM were similar in size and shape to those of the reported virus [8,11]. The MReo2045 isolate exhibited HA only with pig erythrocytes, similar to other MRVs isolated from pigs and dogs [9,11]. The HA of MRV allows sero-surveillance of MRV using hemagglutination inhibition tests with several animal sera. RT-PCR or nested PCR have been used to detect MRV in animal feces [9,10]. RT-PCR amplification of MRV based on the S1 and S4 genes produced bands of the expected size. This suggests that MRV-3 primers may be used for the diagnosis of MRV.
The pathogenicity of MRV in roe deer is unclear, because it is difficult to obtain experimental infection results in wild animals, and naturally occurring MRV infections have been reported in several animal species [4]. A recent study found that mice were easily infected with MRV and developed respiratory disease [7]. Therefore, pathogenic tests are required in experimental animals, such as BALB/c mice, to assess whether the MReo2045 isolate is pathogenic.
The MRV S1 gene has been used to study evolutionary relationships and identify MRV serotypes [7,8]. It has been reported that the S1 gene was responsible for spreading to the hostŌĆÖs central nervous system [21]. When the partial S1 genes of 20 MRVs were compared, the MReo2045 isolate was classified as belonging to serotype 3 and closely related to the T3/Bat/Germany/342/08 and T3D/04 strains, which were isolated from bats and dogs. These results suggest that the roe deer MRV originated via the reassortment of MRVs from other wild animals, including bats.
The partial amino-acid sequences of the Žā1 protein were aligned, which revealed respective 10 and 37 amino-acid differences between the MReo2045 isolate and the T3/Bat/Germany/342/08 and KPR G strains. Therefore, the MReo2045 isolate may be distantly related to the KPR G strain. Because MRV is prevalent in livestock and wild animals, further investigations of MRV in animals are needed. Recently, it was reported that MRV induced cell death preferentially in transformed cells and reovirus T3D mutant could infect human tumor cells [22,23]. It may be also necessary to study MRV as a novel oncolytic agent.
In conclusion, we isolated a virus from a nasal swab of a dead roe deer using Vero cells and confirmed that the isolate was an MRV-3 strain closely related to the T3/Bat/Germany/342/08 strain, which was reported in Germany. The MReo2045 isolate may provide raw material for developing a new diagnostic reagent and could be used as an antigen for serological monitoring.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print