ņä£ļĪĀ
ļÅ╝ņ¦Ćņ£ĀĒ¢ēņä▒ņäżņé¼ļ░öņØ┤ļ¤¼ņŖż(porcine epidemic diarrhea virus, PEDV)ļŖö Nidoviralesļ¬® CoronaviridaeĻ│╝ AlphacoronavirusņåŹņŚÉ ņåŹĒĢśļŖö ņÖĖĒö╝ļź╝ Ļ░Ćņ¦ĆļŖö ļŗ©ņØ╝Ļ░ĆļŗźņØś RNA ļ░öņØ┤ļ¤¼ņŖżļĪ£ņä£ ņøÉĒśĢņØ┤ļ®░ ņÖĖĻ│ĮņŚÉ ņÖĢĻ┤Ćļ¬©ņ¢æņ£╝ļĪ£ ļŗżņłśņØś ņŖżĒīīņØ┤Ēü¼ļź╝ Ļ░Ćņ¦ĆļŖö Ļ▓āņØ┤ ĒŖ╣ņ¦ĢņØ┤ļŗż[1]. PEDVļŖö ļÅ╝ņ¦ĆņĀäņŚ╝ņä▒ņ£äņןņŚ╝ļ░öņØ┤ļ¤¼ņŖż(transmissible gastroenteritis virus, TGEV)ņÖĆ Ļ░ÖņØĆ ņĮöļĪ£ļéśļ░öņØ┤ļ¤¼ņŖżņØ┤ņ¦Ćļ¦ī ĒĢŁņøÉņä▒ņØ┤ ņāüņØ┤ĒĢśļŗż. PEDļŖö 1971ļģä ņśüĻĄŁņŚÉņä£ ņĄ£ņ┤łļĪ£ ļ░£ņāØĒĢśņŚ¼ Ēżņ£Āņ×ÉļÅłņØä ņĀ£ņÖĖĒĢ£ ņ£Īņä▒ ļ╣äņ£ĪļÅłņŚÉņä£ ĻĖēņä▒ ņäżņé¼ņ”ØņØä ļéśĒāĆļé┤ņ¢┤ ĻĖ░ņĪ┤ņŚÉ ņĢīļĀżņĀĖ ņ׳ļŖö TGEņÖĆ ļ¦żņÜ░ ņ£Āņé¼ĒĢ£ ņ”ØņāüņØä ļéśĒāĆļé┤ņŚłļŗż[2]. ņØ┤Ēøä ļŗżļźĖ ņ£Āļ¤Į ĻĄŁĻ░ĆļōżļĪ£ ņĀäĒīīļÉśņ¢┤ ņØ┤ ņ¦łļ│æņØä epidemic viral diarrhea (EVD)ļĪ£ ļ¬ģļ¬ģĒĢśņśĆļŗż. ĻĘĖ Ēøä 1976ļģä ņśüĻĄŁņŚÉņä£ Ēżņ£Āņ×ÉļÅłņØä ĒżĒĢ©ĒĢ£ ļ¬©ļōĀ ņŚ░ļĀ╣ņĖĄņØś ļÅ╝ņ¦ĆņŚÉņä£ ĻĖēņä▒ ņäżņé¼ņ”ØņØ┤ ļ░£ņāØĒĢśņśĆļŗż. ņØ┤ļ¤¼ĒĢ£ ļæÉ ļ▓łņØś ņäżņé¼ņ”Ø ļ░£ņāØ ļÅ╝ņ¦ĆņŚÉņä£ TGEVņÖĆ ĻĖ░ĒāĆ ņןļ│æņøÉņä▒ ņøÉņØĖņ▓┤ļŖö Ļ▓ĆņČ£ļÉśņ¦Ć ņĢŖņĢśņ£╝ļ®░, ļæÉ ņ¦łļ│æņØä ĻĄ¼ļČäĒĢśĻĖ░ ņ£äĒĢśņŚ¼ 1971ļģä ļ░£ņāØĒĢ£ ņäżņé¼ņ”ØņØĆ EVD 1ĒśĢņ£╝ļĪ£, 1976ļģä ļ░£ņāØ ņäżņé¼ņ”ØņØĆ EVD 2ĒśĢņ£╝ļĪ£ ļ¬ģļ¬ģĒĢśņśĆļŗż. ņØ┤Ēøä EVD 2 ļ░£ņāØ ļÅ╝ņ¦ĆņŚÉņä£ ņĮöļĪ£ļéśļ░öņØ┤ļ¤¼ņŖż ņ£Āņé¼ ļ│æņøÉņ▓┤Ļ░Ć ļČäļ”¼ļÉśņŚłĻ│Ā, ņØ┤ ļ░öņØ┤ļ¤¼ņŖżĻ░Ć EVD 1 ļ░£ņāØņŚÉļÅä Ļ┤ĆņŚ¼ĒĢśņśĆļŹś ņĀÉņØ┤ ļ░ØĒśĆņĀĖ ņØ┤ ņäżņé¼ļ│æņØä PEDļĪ£ ļ¬ģļ¬ģĒĢĀ Ļ▓āņØä ņĀ£ņĢłĒĢśņśĆņ£╝ļ®░, Ēśäņ×¼Ļ╣īņ¦Ć ņĀä ņäĖĻ│äņĀüņ£╝ļĪ£ ĒåĄņÜ®ļÉśĻ│Ā ņ׳ļŗż[2].
ņĢäņŗ£ņĢäņŚÉņä£ļŖö ņØ╝ļ│ĖņŚÉņä£ 1983ļģä ļ░£ņāØ ļ│┤Ļ│ĀĻ░Ć ņ׳ņŚłņ£╝ļ®░[3], ĻĄŁļé┤ņŚÉņä£ļŖö 1992ļģä ņäżņé¼ ņ”ØņāüņØä ļ│┤ņØ┤ļŖö ņ×ÉļÅłņ£╝ļĪ£ļČĆĒä░ PEDVļź╝ ņ▓śņØīņ£╝ļĪ£ ļČäļ”¼ĒĢśņśĆļŗż[4]. ņłśņØśĻ│╝ĒĢÖņŚ░ĻĄ¼ņåī(Ēśä ļåŹļ”╝ņČĢņé░Ļ▓ĆņŚŁļ│ĖļČĆ)ņØś ļ│┤Ļ│ĀņŚÉ ļö░ļź┤ļ®┤, 1992ļģä 1ņøöļČĆĒä░ 1993ļģä 12ņøöĻ╣īņ¦Ć ņØśļó░ļÉ£ 71Ļ▒┤ņØś ļ░öņØ┤ļ¤¼ņŖżņä▒ ņäżņé¼ ņśł ņżæ 56.3%Ļ░Ć PEDļĪ£ ņ¦äļŗ©ļÉśņŚłņ£╝ļ®░, ĻĘĖ ņżæ 90% ņĀĢļÅäĻ░Ć 10ņØ╝ļĀ╣ ņØ┤ĒĢśņØś Ēżņ£Āņ×ÉļÅłņŚÉ ņ¦æņżæļÉśņ¢┤ ņ׳ņŚłļŗż[5]. ĻĘĖ ņØ┤ĒøäņŚÉļŖö ņĀäĻĄŁņĀüņ£╝ļĪ£ ĒÖĢņé░ļÉśņ¢┤ ņ¦ĆņåŹņĀüņ£╝ļĪ£ ļ░£ņāØĒĢśņśĆņ£╝ļéś 2011-2012ļģä ņé¼ņØ┤ņŚÉļŖö ļŗżņåī ņåīĻ░ĢņāüĒā£ļź╝ ļéśĒāĆļé┤ņŚłļŗż. ĻĘĖļ¤¼ļéś 2013ļģäļČĆĒä░ ņĀäĻĄŁ ņ¢æļÅł ļåŹĻ░ĆņŚÉņä£ PEDĻ░Ć ņ×¼ļ░£ĒĢśĻ│Ā ļ╣Āļź┤Ļ▓ī ĒÖĢņé░ļÉśņ¢┤ Ēü░ Ļ▓ĮņĀ£ņĀü Ēö╝ĒĢ┤ļź╝ ņŻ╝Ļ│Ā ņ׳ļŗż[6,7]. ņĀ£ņŻ╝ņŚÉņä£ļŖö PEDĻ░Ć 2004ļģä 3ņøö ļ¦łņ¦Ćļ¦ēņ£╝ļĪ£ ļ░£ņāØĒĢ£ Ēøä ņĢĮ 10ļģäĻ░ä ļ»Ėļ░£ņāØņØ┤ņŚłņ£╝ļéś, 2014ļģä 4ņøöļČĆĒä░ ļŗżņŗ£ ļ░£ņāØĒĢśņŚ¼ Ēśäņ×¼Ļ╣īņ¦Ć ņ¢æļÅł ņé░ņŚģņŚÉ Ēü░ ĒāĆĻ▓®ņØä ņŻ╝Ļ│Ā ņ׳ļŖö ņŗżņĀĢņØ┤ļŗż.
ļö░ļØ╝ņä£ ļ│Ė ņŚ░ĻĄ¼ļŖö ņĀ£ņŻ╝ņ¦ĆņŚŁņØś ņ×¼ļ░£ņāØ ņŗ£ņĀÉņØĖ 2014ļģä 4ņøöļČĆĒä░ 18Ļ░£ņøö ļÅÖņĢłņØś PED ļ░£ņāØ ņāüĒÖ®ņØä ļČäņäØĒĢśĻ│Ā, Ļ░ÉņŚ╝ļÉ£ ļÅ╝ņ¦ĆņŚÉ ļīĆĒĢ£ ļ│æļ”¼ĒĢÖņĀü Ļ▓Ćņé¼ņÖĆ ĒĢ©Ļ╗ś ļČäļ”¼ļÉ£ ļ░öņØ┤ļ¤¼ņŖżņŚÉ ļīĆĒĢ£ ņ£ĀņĀäĒĢÖņĀü ļČäņäØņØä ņŗżņŗ£ĒĢśņśĆļŗż. ļśÉĒĢ£ ņżæĻĄŁĻ│╝ ļ»ĖĻĄŁņŚÉņä£ ļ░£ņāØĒĢśņśĆļŹś PEDVņÖĆ ņĀ£ņŻ╝ņŚÉņä£ ļČäļ”¼ļÉ£ ļ░öņØ┤ļ¤¼ņŖżņØś ņ£ĀņĀäĒśĢņØä ļ╣äĻĄÉĒĢśĻ│Āņ×É ĒĢśņśĆļŗż.
ņ×¼ļŻī ļ░Å ļ░®ļ▓Ģ
Ļ│Ąņŗ£ņ×¼ļŻī ļ░Å PED ļ░£ņāØ ĒśäĒÖ® ņĪ░ņé¼
Ļ│Ąņŗ£ļÅÖļ¼╝ņØĆ 2014ļģä 4ņøöļČĆĒä░ 2015ļģä 9ņøöĻ╣īņ¦Ć ņĀ£ņŻ╝ļÅä ļé┤ ņ¢æļÅł ļåŹĻ░Ć 52ĒśĖņŚÉņä£ 72ĒÜīņŚÉ Ļ▒Ėņ│É ņĀ£ņŻ╝ĒŖ╣ļ│äņ×Éņ╣śļÅä ļÅÖļ¼╝ņ£äņāØņŗ£ĒŚśņåīņŚÉ ļ│æņä▒Ļ░ÉņĀĢ ņØśļó░ļÉ£ ļÅ╝ņ¦Ć 153ļ¦łļ”¼ļź╝ ļīĆņāüņ£╝ļĪ£ ĒĢśņśĆļŗż. ļåŹĻ░ĆņŚÉņä£ ņØśļó░ļÉ£ ļÅ╝ņ¦ĆņŚÉ ļīĆĒĢ£ PED ņ¦äļŗ©ņØĆ ņŚŁņĀäņé¼ ņżæĒĢ®ĒÜ©ņåīņŚ░ņćäļ░śņØæ(reverse transcription polymerase chain reaction, RT-PCR) Ļ▓Ćņé¼ Ļ▓░Ļ│╝ļź╝ ĒåĀļīĆļĪ£ ņĄ£ņóģ ĒÖĢņ¦äĒĢśņśĆļŗż. PEDļĪ£ ņ¦äļŗ©ļÉ£ ļÅ╝ņ¦Ćļź╝ ņŚ░ļĀ╣ļ│ä, Ļ│äņĀłļ│ä ļ░Å ņ¦ĆņŚŁļ│äļĪ£ ĻĄ¼ļČäĒĢśņŚ¼ ļ░£ņāØ ņāüĒÖ®ņØä ļČäņäØĒĢśņśĆļŗż.
ņ£ĪņĢłĻ▓Ćņé¼
ļÅÖļ¼╝ņ£äņāØņŗ£ĒŚśņåīņŚÉ ņØśļó░ļÉ£ ļÅ╝ņ¦ĆņŚÉ ļīĆĒĢśņŚ¼ ņ×äņāüņ”ØņāüņØä ĒÖĢņØĖĒĢśĻ│Ā, ņÖĖĻ┤Ć Ļ▓Ćņé¼ Ēøä ņØ╝ļ░śņĀüņØĖ ļČĆĻ▓ĆņłĀņŗØņŚÉ ļö░ļØ╝ņä£ ļé┤ļČĆņןĻĖ░ņØś ļ│æļ│ĆņØä Ļ┤Ćņ░░ĒĢśņśĆļŗż. ĒŖ╣Ē׳ ņ£äņןĻ┤ĆņØä ĒżĒĢ©ĒĢ£ ņåīĒÖöĻĖ░Ļ│äņØś ļ│ĆĒÖöņāüņØä ļ®┤ļ░ĆĒ׳ Ļ┤Ćņ░░ĒĢśņśĆļŗż. ņØśļó░ļÉ£ ļÅ╝ņ¦ĆņØś ņ£ĪņĢł ļ│æļ│Ć ļ░£ņāØ ļ╣łļÅäļź╝ Ēżņ£Ā ņŚ░ļĀ╣ņŚÉ ļö░ļØ╝ 4ņØ╝ļĀ╣ ļ»Ėļ¦ī, 4ņØ╝ ņØ┤ņāü 7ņØ╝ ņØ┤ĒĢś, 8ņØ╝ ņØ┤ņāüņ£╝ļĪ£ ņäĖļČäĒĢśņŚ¼ ļ╣äĻĄÉĒĢśņśĆļŗż.
ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀü Ļ▓Ćņé¼
ļÅ╝ņ¦Ć ļČĆĻ▓Ć ņŗ£ ņŻ╝ņÜö ļé┤ļČĆ ņŗżņ¦ł ņןĻĖ░ļź╝ ņ▒äņĘ©ĒĢśņśĆļŗż. ĒŖ╣Ē׳, ņåīĒÖöĻĖ░Ļ│äļŖö ņ£ä, ņåīņן(ņŗŁņØ┤ņ¦Ćņן, Ļ│Ąņן ļ░Å ĒÜīņן), ļīĆņן(ļ¦╣ņן ļ░Å Ļ▓░ņן)ņØä ĻĄ¼ļČäĒĢśņŚ¼ ņ▒äņĘ©ĒĢ£ ļŗżņØī 10% ņżæņä▒ņÖäņČ®Ēżļź┤ļ¦Éļ”░ņŚÉ Ļ│ĀņĀĢĒĢśņŚ¼ ņØ╝ļ░śņĀüņØĖ ņĪ░ņ¦üņ▓śļ”¼ ļ░®ļ▓ĢņŚÉ ļö░ļØ╝ ĒīīļØ╝ĒĢĆ Ēżļ¦żĒĢśņŚ¼ 3-4 ╬╝m ļæÉĻ╗śļĪ£ ņĀłĒÄĖņØä ņĀ£ņ×æĒĢ£ Ēøä hematoxylinĻ│╝ eosin ņŚ╝ņāēņØä ņŗżņŗ£ĒĢśņŚ¼ Ļ┤æĒĢÖĒśäļ»ĖĻ▓Įņ£╝ļĪ£ Ļ▓ĆĻ▓ĮĒĢśņśĆļŗż.
ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀü Ļ▓Ćņé¼Ļ░Ć ņłśĒ¢ēļÉ£ ņ┤Ø 153ļ¦łļ”¼ņØś ļÅ╝ņ¦Ć ņżæ ņé¼Ēøäļ│ĆĒÖöņÖĆ ņØĖņ£äņĀü ņåÉņāü(artifact)ņØ┤ ņłśļ░śļÉśņ¦Ć ņĢŖņØĆ 124ļ¦łļ”¼ņØś ņåīņן ņĪ░ņ¦üņŚÉņä£ ņ£Ąļ¬©ņØś ņ£äņČĢ ņĀĢļÅäļź╝ ņłśņ╣śĒÖöĒĢśĻĖ░ ņ£äĒĢśņŚ¼ ņ£Ąļ¬©ņØś ļåÆņØ┤(villous height, VH)ņÖĆ ņøĆņØś Ļ╣ŖņØ┤(crypt depth, CD)ļź╝ ļ╣äĻĄÉĒĢśņśĆļŗż. ĒŖ╣Ē׳ Ļ┤æĒĢÖĒśäļ»ĖĻ▓Į 40ļ░░ ņŗ£ņĢ╝ņŚÉņä£ ļ¼┤ņ×æņ£äļĪ£ ņäĀņĀĢĒĢ£ Ļ│ĄņןĻ│╝ ĒÜīņן, Ļ░ü 10Ļ░£ ļČĆņ£äļź╝ ņĖĪņĀĢĒĢśņŚ¼ ĒÅēĻĘĀ ļ░Å Ēæ£ņżĆĒÄĖņ░©ļź╝ ņé░ņČ£ĒĢśņśĆņ£╝ļ®░, ņØ┤ļź╝ ĒåĀļīĆļĪ£ VHņÖĆ CDņØś ļ╣äņ£©ņØä ņé░ņĀĢĒĢśņśĆļŗż. ļśÉĒĢ£ ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝ņ£╝ļĪ£ ņØĖĒĢ┤ ņåÉņāü ļ░øņØĆ ņ£Ąļ¬©Ļ░Ć ņä£ļĪ£ ņ£ĄĒĢ®ĒĢśļŖö ņĀĢļÅäļź╝ ņäĖļČäĒĢśņŚ¼ ļČäļźśĒĢśņśĆļŗż. Ļ│Ąņן ļ░Å ĒÜīņןņØä Ļ┤æĒĢÖĒśäļ»ĖĻ▓Įņ£╝ļĪ£ Ļ┤Ćņ░░ĒĢśņśĆņØä ļĢī, 100ļ░░ ņŗ£ņĢ╝ņŚÉņä£ ņ£Ąļ¬© ņ£ĄĒĢ®ņØś ņĀĢļÅäĻ░Ć 10% ļ»Ėļ¦īņØ╝ Ļ▓ĮņÜ░ focal (G1), 10-50%ļŖö multifocal (G2), 50% ņØ┤ņāüņØĆ diffuse (G3)ļĪ£ ĻĄ¼ļČäĒĢśņśĆļŗż.
ņ┤Ø 153ļ¦łļ”¼ ņżæ ņé¼Ēøäļ│ĆĒÖöņÖĆ ņØĖņ£äņĀü ņāüĒö╝ ĒāłļØĮņØ┤ ļÅÖļ░śļÉśņ¢┤ ņ׳ņ¦Ć ņĢŖņØĆ Ļ░£ņ▓┤ņØĖ 126ļ¦łļ”¼ņØś Ļ│ĄņןĻ│╝ 120ļ¦łļ”¼ņØś ĒÜīņןņØä ļīĆņāüņ£╝ļĪ£ ņåīņן ņĀÉļ¦ēņāüĒö╝ņäĖĒżņØś ļ│ĆĒÖöņāüņØä ņóĆ ļŹö ļ®┤ļ░ĆĒ׳ Ļ┤Ćņ░░ĒĢśĻ│Ā ņäĖļČäĒĢśņŚ¼ ĻĖ░ļĪØĒĢśņśĆļŗż. Ļ┤æĒĢÖĒśäļ»ĖĻ▓Į 100ļ░░ ļśÉļŖö 200ļ░░ ņŗ£ņĢ╝ņŚÉņä£ ņ£Ąļ¬©ņāüĒö╝ņäĖĒżņØś ĒāłļØĮ(exfoliation), ĒÄĖĒÅēĒÖö(squamation) ļ░Å ņäĖĒżņ¦łņØś Ļ│ĄĒżĒÖö(vacuolation)ņØś ņĀĢļÅäņŚÉ ļö░ļØ╝ 10% ļ»Ėļ¦īņØĖ Ļ▓ĮņÜ░ focal (G1), 10-50%ļŖö multifocal (G2), 50% ņØ┤ņāüņØĆ diffuse (G3)ļĪ£ ĻĄ¼ļČäĒĢśņśĆļŗż. ļśÉĒĢ£ ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀü ļ│æļ│ĆļÅä ņ£ĪņĢł ļ│æļ│ĆĻ│╝ ļ¦łņ░¼Ļ░Ćņ¦ĆļĪ£ ļÅ╝ņ¦ĆņØś ņŚ░ļĀ╣ņŚÉ ļö░ļØ╝ 4ņØ╝ļĀ╣ ļ»Ėļ¦ī, 4ņØ╝ ņØ┤ņāü 7ņØ╝ ņØ┤ĒĢś, 8ņØ╝ ņØ┤ņāüņ£╝ļĪ£ ņäĖļČäĒĢśņŚ¼ ļ░£ņāØ ļ╣łļÅäļź╝ ļ╣äĻĄÉĒĢśņśĆļŗż.
ļ®┤ņŚŁņĪ░ņ¦üĒÖöĒĢÖ Ļ▓Ćņé¼
ņØśļó░ļÉ£ ļÅ╝ņ¦Ć ņŗ£ļŻī ņżæ ņŗ¼ĒĢ£ ņé¼Ēøäļ│ĆĒÖöĻ░Ć ņ¦äĒ¢ēļÉ£ Ļ▓āņØä ņĀ£ņÖĖĒĢśĻ│Ā 135ļ¦łļ”¼ņØś Ļ│Ąņן, 127ļ¦łļ”¼ņØś ĒÜīņן ļ░Å 133ļ¦łļ”¼ņØś Ļ▓░ņןņŚÉņä£ ļīĆĒĢśņŚ¼ PEDV ĒĢŁņøÉņØś ļČäĒżļź╝ ĒÖĢņØĖĒĢśĻĖ░ ņ£äĒĢśņŚ¼ ļ®┤ņŚŁņĪ░ņ¦üĒÖöĒĢÖ(immunohistochemistry, IHC) Ļ▓Ćņé¼ļź╝ ņłśĒ¢ēĒĢśņśĆļŗż. ĒīīļØ╝ĒĢĆ ņĪ░ņ¦üņØä 4-5 ╬╝m ļæÉĻ╗śļĪ£ ļ░ĢņĀłĒĢśņŚ¼ silane ņĮöĒīģ ņŖ¼ļØ╝ņØ┤ļō£ņŚÉ ļČĆņ░®ĒĢśĻ│Ā ĒāłĒīīļØ╝ĒĢĆ ļ░Å ĒĢ©ņłśĻ│╝ņĀĢņØä Ļ▒░ņ│żļŗż. ņĪ░ņ¦ü ņŖ¼ļØ╝ņØ┤ļō£ļź╝ 3% H2O2Ļ░Ć ņ▓©Ļ░ĆļÉ£ phosphate buffered saline (PBS, pH 7.2)ņŚÉ 10ļČäĻ░ä ļ░śņØæĒĢśĻ│Ā, ĒĢŁņøÉņä▒ņØś ļČĆĒÖ£ņØä ņ£äĒĢ┤ citrate buffer solutionņ£╝ļĪ£ 97ŌäāņŚÉņä£ 60ļČäĻ░ä ļ░śņØæĒĢśņśĆļŗż. 1ņ░© ĒĢŁņ▓┤ļĪ£ļŖö mouse monoclonal anti-PEDV antibody (Median Diagnostics, Korea)ļź╝ 1:500ļ░░ļĪ£ ĒؼņäØĒĢ£ ļŗżņØī ņĪ░ņ¦üņĀłĒÄĖņŚÉ ņĀüĒĢśĒĢśņŚ¼ 37ŌäāņŚÉņä£ 1ņŗ£Ļ░ä ļ░śņØæĒĢśņśĆļŗż. 2ņ░© ĒĢŁņ▓┤ļĪ£ EnVision/horseradish peroxidase, rabbit/mouse reagent (Dako, Denmark)ļź╝ ņŖ¼ļØ╝ņØ┤ļō£ņŚÉ ņĀüĒĢśĒĢśņŚ¼ 37ŌäāņŚÉņä£ 40ļČäĻ░ä ļ░śņØæĒĢśņśĆļŗż. Ļ░ü ļŗ©Ļ│äļ│äļĪ£ ļ░śņØæ ĒøäņŚÉļŖö PBSļĪ£ 5ļČäņö® 2ĒÜīņŚÉ Ļ▒Ėņ│É ņłśņäĖĒĢśņśĆļŗż. ļ¬©ļōĀ ļ░śņØæņØ┤ ļüØļé£ ņĀłĒÄĖņØĆ 3,3ŌĆś-diamino-benzidine tetrahydrochloride (Dako)ļĪ£ ļ░£ņāēņŗ£Ēé© Ēøä, 3ņ░© ņ”ØļźśņłśņŚÉņä£ ļ░śņØæņØä ņóģĻ▓░ĒĢśņśĆļŗż. ļīĆņĪ░ņŚ╝ņāēņØĆ Mayer's hematoxylin (Sigma, USA)ņ£╝ļĪ£ ņŚ╝ņāēĒĢśņśĆņ£╝ļ®░, ĒāłņłśņÖĆ Ēł¼ļ¬ģĻ│╝ņĀĢņØä Ļ▒░ņ│É ļ┤ēņ×ģĒĢśņśĆļŗż.
ļ®┤ņŚŁņŚ╝ņāēņØ┤ ļüØļé£ ņĪ░ņ¦ü ļé┤ PEDV ĒĢŁņøÉņØś ļ░£Ēśä ņĀĢļÅäļŖö ĻĖ░ņĪ┤ņØś ļ│┤Ļ│Āļź╝ ĒåĀļīĆļĪ£ 3ļō▒ĻĖē(expression grade, EG)ņ£╝ļĪ£ ņäĖļČäĒĢśņŚ¼ ĒÅēĻ░ĆĒĢśņśĆļŗż[8]. ņ”ē, Ļ┤æĒĢÖĒśäļ»ĖĻ▓Į 200ļ░░ ņŗ£ņĢ╝ņŚÉņä£ Ļ│Ąņן, ĒÜīņן ļ░Å Ļ▓░ņן ņĀÉļ¦ēņāüĒö╝ņäĖĒżņŚÉņä£ PEDV ĒĢŁņøÉņØ┤ ņĀäņ▓┤ ņāüĒö╝ņäĖĒżņØś 10% ļ»Ėļ¦īņŚÉņä£ ļ░£ĒśäļÉśļŖö Ļ▓ĮņÜ░ ĻĄŁņåīņĀü(focal, EG1), 10-50%ļŖö ļŗżļ│æņåīņä▒(multifocal, EG2), 50% ņØ┤ņāüņØĆ ļ»Ėļ¦īņä▒(diffuse, EG3)ņ£╝ļĪ£ ĻĄ¼ļČäĒĢśņśĆļŗż.
ļ│æņøÉņ▓┤ Ļ▓Ćņé¼
Ļ▓Ćņé¼ņŗ£ļŻīņØś RNA ņČöņČ£
ņäżņé¼ ņ£Āļ░£ ļ│æņøÉņ▓┤ļź╝ Ļ▓Ćņé¼ĒĢśĻĖ░ ņ£äĒĢśņŚ¼ ņ┤Ø 153ļæÉņØś ļÅ╝ņ¦Ć ļīĆņןņŚÉņä£ ļ¼┤ĻĘĀņĀüņ£╝ļĪ£ ļČäļ│ĆņØä ņ▒äņĘ© Ēøä -70ŌäāņŚÉ ļ│┤Ļ┤ĆĒĢśņŚ¼ ļ░öņØ┤ļ¤¼ņŖżĻ▓Ćņé¼ņŚÉ ĒÖ£ņÜ®ĒĢśņśĆļŗż. RNA ļ░öņØ┤ļ¤¼ņŖżņØĖ PEDV ļ░Å TGEVņŚÉ ļīĆĒĢ£ RT-PCRņØä ņŗżņŗ£ĒĢśņśĆļŗż. ņ£ĀņĀäņ×ÉĻ▓Ćņé¼ļź╝ ņ£äĒĢśņŚ¼ ļČäļ│Ćņŗ£ļŻīļź╝ DNase RNase free distilled water (Invitrogen, USA)ņÖĆ 1:10 ļ╣äņ£©ļĪ£ Ēś╝ĒĢ®ĒĢśņŚ¼ ņāüņĖĄņĢĪņØä ņØ┤ņÜ®ĒĢśņśĆļŗż. RNA ņČöņČ£ņØĆ RNeasy Mini kit (QIAGEN GmbH, Germany)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņĀ£ņĪ░ņé¼Ļ░Ć ņĀ£ņŗ£ĒĢ£ ņŗżĒŚśļ░®ļ▓ĢņŚÉ ņżĆĒĢśņŚ¼ ņŗżņŗ£ĒĢśņśĆļŗż.
RT-PCR ļ░śņØæ ņĪ░Ļ▒┤
RT-PCRņØĆ PEDV ļ░Å TGEVļź╝ Ļ▓ĆņČ£ĒĢśĻĖ░ ņ£äĒĢ£ primerĻ░Ć ĒĢ©ņ£ĀļÉśņ¢┤ ņ׳ļŖö ņĀ£ĒÆłĒÖöļÉ£ premixņØĖ i-TGEV/PEDV detection kit (Intronbiotechnology, Korea)ņŚÉ ņČöņČ£ĒĢ£ RNA 2 ╬╝L ļ░Å DNase RNase free distilled water (Invitrogen) 18 ╬╝Lļź╝ ļČäņŻ╝ĒĢśņśĆņ£╝ļ®░, ņĀ£ņŗ£ļÉ£ ņŗżĒŚśļ░®ļ▓ĢņŚÉ ņżĆĒĢśņŚ¼ ņłśĒ¢ēĒĢśņśĆļŗż.
RT-PCR ņ”ØĒÅŁņé░ļ¼╝ņØś ĒÖĢņØĖ
ļ░śņØæ ņóģļŻī Ēøä, Ļ░üĻ░üņØś ļ░śņØæņĢĪ 8 ╬╝Lņö®ņØä 1.5% agarose gel ņāüņŚÉņä£ ņĀäĻĖ░ņśüļÅÖņØä ņŗżņŗ£ĒĢ£ ļŗżņØī RedSafe nucleic acid staining solution (Intronbiotechnology) ņÜ®ņĢĪ(0.5 ╬╝L/mL in distilled water)ņ£╝ļĪ£ ņŚ╝ņāēĒĢśņśĆļŗż. Ultraviolet transilluminatiorļĪ£ Ļ░üĻ░üņØś ņ£ĀņĀäņ×ÉņŚÉ ļīĆĒĢ£ ĒŖ╣ņØ┤ņĀüņØĖ ļ░┤ļō£ ņ£Āļ¼┤ļź╝ ĒÖĢņØĖĒĢśņśĆļŗż.
PEDVņØś ņ£ĀņĀäĒĢÖņĀü Ļ▓Ćņé¼
ņĀ£ņŻ╝ļÅä ļÅ╝ņ¦Ć ņŗ£ļŻīņŚÉņä£ Ļ▓ĆņČ£ļÉ£ PEDV ņżæ 12ņŻ╝(2014ļģä 8ņŻ╝: KOR/JJ-1-8/2014, 2015ļģä 4ņŻ╝: KOR/JJ-1-4/2015)ņŚÉ ļīĆĒĢ£ ņ£ĀņĀäĒĢÖņĀü Ļ▓Ćņé¼ļź╝ ņŗżņŗ£ĒĢśņśĆļŗż. PEDV spike (S) ņ£ĀņĀäņ×É Ļ▓ĆņČ£ņØä ņ£äĒĢ£ primerļŖö Chen ļō▒[9]ņØś ļ░®ļ▓ĢņŚÉ ļö░ļØ╝ ņĀ£ņ×æĒĢśņśĆļŗż. PEDV RNAļŖö ņäżņé¼ ļČäļ│ĆņØ┤ļéś ņåīņן ņĪ░ņ¦üņ£╝ļĪ£ļČĆĒä░ TRIzol LS (Invitrogen Corp., USA)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņČöņČ£ĒĢśņśĆļŗż. RT-PCRņØĆ RT ļŗ©Ļ│äļŖö 50ŌäāņŚÉņä£ 30ļČä, pre-cycle heating ļŗ©Ļ│äļŖö 95ŌäāņŚÉņä£ 5ļČä, cycle reactions ļŗ©Ļ│äļŖö 94ŌäāņŚÉņä£ 30ņ┤ł, 52ŌäāņŚÉņä£ 2ļČä 30ņ┤ł, 72ŌäāņŚÉņä£ 1ļČä 30ņ┤łĻ░ä 35 cyclesļź╝ ņłśĒ¢ēĒĢ£ Ēøä ņ”ØĒÅŁļÉ£ ņé░ļ¼╝ņØä pGEM-T vector system II (Cat. No. A3610; Promega Corp., USA)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ Ēü┤ļĪ£ļŗØĒĢśņśĆļŗż.
PEDV ņ£ĀņĀäņ×Éļź╝ ļČäņäØĒĢśĻĖ░ ņ£äĒĢśņŚ¼ Ēü┤ļĪ£ļŗØļÉ£ ņ£ĀņĀäņ×Éļź╝ T7Ļ│╝ SP6 sequencing primerļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļ¦łĒü¼ļĪ£ņĀĀ(Macrogen, Korea)ņŚÉ ABI Prism 3730xi DNA sequencerļĪ£ ņ£ĀņĀäņ×É ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ĒīÉļÅģĒĢśņśĆņ£╝ļ®░, ĒīÉļÅģļÉ£ S ņ£ĀņĀäņ×ÉļōżņØä ļéśņŚ┤ĒĢśĻĖ░ ņ£äĒĢ┤ CLUSTALX alignment programĻ│╝ BIOEDIT 7.053 programņØä ņé¼ņÜ®ĒĢśņŚ¼ ļČäņäØĒĢśņśĆļŗż.
ņØ┤Ēøä, ņ£ĀņĀäņ×ÉĻ▓Ćņé¼ Ļ▓░Ļ│╝ļź╝ ĒåĀļīĆļĪ£ Ļ│äĒåĄĒĢÖņĀü ĒŖĖļ”¼ļź╝ ļ¦īļōżĻĖ░ ņ£äĒĢ┤ Mega ver. 6.0 (Mega Software, USA) ļé┤ņŚÉ ļōżņ¢┤ņ׳ļŖö neighbor-joining methodļź╝ ņé¼ņÜ®ĒĢśņśĆļŗż[10].
ĒåĄĻ│ä ņ▓śļ”¼
IHC Ļ▓Ćņé¼ļź╝ ĒåĄĒĢ£ Ļ░£ņ▓┤ļ│ä PEDV ĒĢŁņøÉņØś ļ░£Ēśä ļō▒ĻĖē(EG)ņØĖ EG1, EG2, EG3ļź╝ Ļ░üĻ░ü 1, 2, 3ņ£╝ļĪ£ ņłśņ╣śĒÖöĒĢśņŚ¼ ĒĢ®ĒĢ£ ļŗżņØī ņĀäņ▓┤ Ļ░£ņ▓┤ņłśļĪ£ ļéśļłäņ¢┤ ĒÅēĻĘĀ ĒĢŁņøÉ ļ░£Ēśä ļō▒ĻĖēĻ│╝ Ēæ£ņżĆĒÄĖņ░©ļź╝ ņé░ņČ£ĒĢśņśĆļŗż. ļÅ╝ņ¦Ć Ļ│Ąņן, ĒÜīņן ļ░Å Ļ▓░ņן ņĪ░ņ¦ü ļé┤ PEDV ĒĢŁņøÉņØś ļ░£Ēśä ņĀĢļÅäņŚÉ ņ£ĀņØśņä▒ ņ׳ļŖö ņ░©ņØ┤Ļ░Ć ņ׳ļŖöņ¦Ćļź╝ ĒÖĢņØĖĒĢśĻĖ░ ņ£äĒĢśņŚ¼ IBM SPSS Statistics ver. 25.0 (IBM Corp., USA)ņØä ņé¼ņÜ®ĒĢśņŚ¼ ANOVA testļĪ£ ĒåĄĻ│äļČäņäØņØä ņŗżņŗ£ĒĢśņśĆņ£╝ļ®░, p < 0.05ņØ╝ ļĢī ņ£ĀņØśĒĢ£ ņ░©ņØ┤Ļ░Ć ņ׳ļŖö Ļ▓āņ£╝ļĪ£ Ļ░äņŻ╝ĒĢśņśĆļŗż.
Ļ▓░Ļ│╝
ņĀ£ņŻ╝ļÅä ļé┤ PED ņ¦äļŗ© ĒśäĒÖ®
ņĀ£ņŻ╝ļÅäņŚÉņä£ 2014ļģä 4ņøöļČĆĒä░ 18Ļ░£ņøö ļÅÖņĢł PEDļĪ£ ņ¦äļŗ©ļÉ£ ņśłļŖö ņ┤Ø 153ļ¦łļ”¼ļĪ£ ĒÖĢņØĖļÉśņŚłļŗż. ļÅ╝ņ¦ĆņØś ņŚ░ļĀ╣ļ│äļĪ£ļŖö 4ņØ╝ļĀ╣ ļ»Ėļ¦īņØ┤ 101ļ¦łļ”¼(66.0%), 4-7ņØ╝ļĀ╣ 43ļ¦łļ”¼(28.1%) ļ░Å 8ņØ╝ļĀ╣ ņØ┤ņāü 9ļ¦łļ”¼(5.9%)ļĪ£ ĒÖĢņØĖļÉśņŚłļŗż. ņØ┤ ņżæ, 8ņØ╝ļĀ╣ ņØ┤ņāü ļÅ╝ņ¦ĆļŖö 9ņØ╝ļĀ╣ 2ļ¦łļ”¼, 10ņØ╝ļĀ╣, 14ņØ╝ļĀ╣, 15ņØ╝ļĀ╣, ļ░Å 17ņØ╝ļĀ╣ņØ┤ Ļ░üĻ░ü 1ļ¦łļ”¼, 21ļĀ╣ 3ļ¦łļ”¼ļĪ£ ĻĄ¼ļČäļÉśņŚłļŗż.
ņĀ£ņŻ╝ĒŖ╣ļ│äņ×Éņ╣śļÅä Ļ░ĆņČĢĒåĄĻ│äņĪ░ņé¼ ņ×ÉļŻīņŚÉ ļö░ļź┤ļ®┤, 2014ļģä Ēśäņ×¼Ļ╣īņ¦Ć ņĀ£ņŻ╝ļÅä ļé┤ ņĀäņ▓┤ ņ¢æļÅł ļåŹĻ░ĆņłśļŖö 304ĒśĖņØ┤ļ®░, Ē¢ēņĀĢņŗ£ ļ│äļĪ£ļŖö ņĀ£ņŻ╝ņŗ£ 214ĒśĖ, ņä£ĻĘĆĒżņŗ£ 90ĒśĖļĪ£ ļéśĒāĆļéś ņ׳ļŗż. PEDĻ░Ć ņ¦äļŗ©ļÉ£ ņ¢æļÅł ļåŹĻ░ĆļŖö ņ┤Ø 52ĒśĖļĪ£ ņĀäņ▓┤ ļåŹĻ░Ć ņżæ ņĢĮ 17.1%ņŚÉ ļŗ¼ĒĢśņśĆļŗż. ņ¦ĆņŚŁļ│äļĪ£ļŖö ņĀ£ņŻ╝ņŗ£ 19.6% (42/214), ņä£ĻĘĆĒżņŗ£ 11.1% (10/90)ļĪ£ ļéśĒāĆļéś ņĀ£ņŻ╝ņŗ£ņŚÉņä£ņØś PED ļ░£ņāØņØ┤ ļŹöņÜ▒ ļåÆņØĆ Ļ▓āņ£╝ļĪ£ ĒÖĢņØĖļÉśņŚłļŗż. ļ░£ņāØ ļåŹĻ░Ćļź╝ ņØŹ, ļ®┤ļ│äļĪ£ ņäĖļČäĒĢśņŚ¼ ņé┤ĒÄ┤ļ│┤ņĢśņØä ļĢī, ņĀ£ņŻ╝ļÅäņØś ņä£ļČĆņ¦ĆņŚŁņŚÉ ĒĢ┤ļŗ╣ĒĢśļŖö ĒĢ£ļ”╝ņØŹ 29.9% (41/137), ļīĆņĀĢņØŹ 27.8% (10/36), ĒĢ£Ļ▓Įļ®┤ 4.2% (1/24)ņŚÉ ĻĄŁĒĢ£ļÉśņ¢┤ ļ░£ņāØĒĢśĻ│Ā ņ׳ņŚłļŗż.
Ļ▓Ćņé¼ ĻĖ░Ļ░ä ļÅÖņĢł ļÅÖņØ╝ĒĢ£ ļåŹņןņŚÉņä£ ņŚ¼ļ¤¼ ņ░©ļĪĆ Ļ▓Ćņé¼ ņØśļó░ļÉ£ Ļ▓ĮņÜ░ļź╝ ĒżĒĢ©ĒĢśņŚ¼ PEDļĪ£ ņ¦äļŗ©ļÉ£ ņśłļŖö ņ┤Ø 72Ļ▒┤(153ļ¦łļ”¼)ņØ┤ņŚłļŗż. ņØ┤ļź╝ ņøöļ│äļĪ£ ņäĖļČäĒĢśņŚ¼ PED ļ░£ņāØņØä ļČäņäØĒĢ£ Ļ▓░Ļ│╝, 2014ļģä 4ņøö 13Ļ▒┤(18.1%), 5ņøö 14Ļ▒┤(19.4%), 6ņøöĻ│╝ 7ņøöņØ┤ Ļ░üĻ░ü 1Ļ▒┤(1.4%), 8ņøö 3Ļ▒┤(4.2%), 9ņøö 4Ļ▒┤(5.6%), 10ņøö 1Ļ▒┤(1.4%), 11ņøöĻ│╝ 12ņøö 2Ļ▒┤(2.8%), 2015ļģä 1ņøö 9Ļ▒┤(12.5%), 2ņøö 5Ļ▒┤(6.9%), 3ņøö 6Ļ▒┤(8.3%), 4ņøö 1Ļ▒┤(1.4%), 5ņøö 3Ļ▒┤(4.2%), 6ņøöĻ│╝ 7ņøö 1Ļ▒┤(1.4%), 9ņøö 5Ļ▒┤(6.9%)ņ£╝ļĪ£ ļéśĒāĆļé¼ļŗż(Table 1). ļö░ļØ╝ņä£ ņĀ£ņŻ╝ļÅä ļé┤ ņĄ£ņ┤ł ņ×¼ļ░£ņāØ ņŗ£ņĀÉņØĖ 2014ļģä 4ņøöĻ│╝ 5ņøöņŚÉ ĒÅŁļ░£ņĀüņØĖ ļ░£ņāØņØä ļ│┤ņØ┤ļŗżĻ░Ć ĒĢśņĀłĻĖ░ņŚÉ ņåīĻ░ĢņāüĒā£ļĪ£ ņĀæņ¢┤ļōżņŚłņ£╝ļéś, ņØ┤Ēøä ļÅÖņĀłĻĖ░ņŚÉ ļōżņ¢┤ņä£ļ®┤ņä£ ļŗżņŗ£ ņ”ØĻ░ĆĒĢśļŖö ņ¢æņāüņØä ļ│┤ņØ┤Ļ│Ā ņ׳ņŚłļŗż.
ĒÖĢņ¦äļÉ£ 52ĒśĖ ļåŹĻ░Ćļź╝ ņØśļó░ ņ£ĀĒśĢļ│äļĪ£ ļČäņäØĒĢ£ Ļ▓░Ļ│╝ 1ĒÜī Ļ▓Ćņé¼ ņØśļó░ ļåŹĻ░ĆļŖö 35ļåŹĻ░Ć(67.3%)ņśĆņ£╝ļ®░, 2ĒÜī, 3ĒÜī ļ░Å 4ĒÜī ņØśļó░ ļåŹĻ░ĆļŖö Ļ░üĻ░ü 15ļåŹĻ░Ć(28.8%), 1ļåŹĻ░Ć(1.9%) ļ░Å 1ļåŹĻ░Ć(1.9%)ļĪ£ ļéśĒāĆļé¼ļŗż.
ņ£ĪņĢłĻ▓Ćņé¼ Ļ▓░Ļ│╝
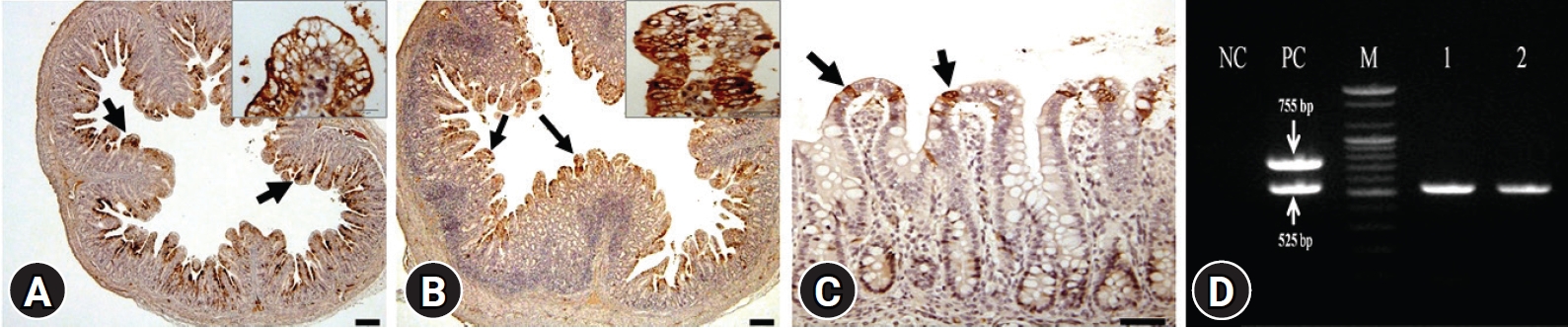
ņ£ĪņĢłņĀüņ£╝ļĪ£ PEDļĪ£ ĒÖĢņ¦äļÉ£ ļÅ╝ņ¦ĆļŖö ĒśäņĀĆĒĢśĻ▓ī ņ£äņČĢļÉśņ¢┤ ļŖæĻ│©ņØ┤ ļģĖņČ£ļÉśĻ│Ā, ņØ╝ļČĆ Ļ░£ņ▓┤ļŖö ņĢłĻĄ¼Ļ░Ć ĒĢ©ļ¬░ļÉśņ¢┤ ņ׳ņŚłļŗż. ĒĢŁļ¼Ė ņŻ╝ņ£ä Ēö╝ļČĆņŚÉļŖö ļŗ┤ĒÖ®ņāē ļśÉļŖö ĒÖ®ņāēņØś ņłśņ¢æņä▒ ļ│Ćļé┤ņÜ®ļ¼╝ņØ┤ ļČĆņ░®ļÉśņ¢┤ ņ׳ņŚłļŗż(Fig. 1A). ņ£ä ļé┤ņŚÉļŖö ņåīĒÖöļÉśņ¦Ć ņĢŖņØĆ ņ£Āļ░▒ņāēņØś ņØæņ£ĀĻ░Ć ļŗżļ¤ē ļōżņ¢┤ņ׳ņŚłļŗż. ņåīņןņØĆ ļŗ┤ņāēņĪ░ļĪ£ Ēć┤ņāēļÉśņ¢┤ ņ׳Ļ▒░ļéś ļ¼┤ļĀźĒĢśņśĆņ£╝ļ®░, ņןļ▓ĮņØ┤ ĒśäņĀĆĒĢśĻ▓ī ņ¢ćņĢäņĀĖ ņןĻ░Ģ ļé┤ņØś ņłśņ¢æņä▒ ļé┤ņÜ®ļ¼╝ņØ┤ ļ¬ģļŻīĒĢśĻ▓ī Ļ┤Ćņ░░ļÉśņŚłļŗż(Fig. 1B). ņåīņן ņןļ¦ēĻ│╝ ņןĻ░äļ¦ēņØś ĒśłĻ┤Ć ļ░Å ļ”╝ĒöäĻ┤ĆņØś Ļ┤Ćņ░░ņØĆ ņÜ®ņØ┤ĒĢśņ¦Ć ņĢŖņĢśļŗż. ņāüļŗ╣ņłś Ļ░£ņ▓┤ņØś ļīĆņן ļé┤Ļ░ĢņŚÉļÅä ĒÖ®ņāēņØś ņłśņ¢æņä▒ ļé┤ņÜ®ļ¼╝ņØ┤ ņČ®ļ¦īļÉśņ¢┤ ņ׳ņŚłļŗż. ņŗĀņןņØś ļŗ©ļ®┤ņāüņŚÉņä£ļŖö ņ£ĀļæÉļČĆ ļśÉļŖö ņŗĀņÜ░ ļČĆņ£äņŚÉ ĒÖ®Ļ░łņāēņØś ņÜöņé░ņŚ╝ņØ┤ ņ╣©ņ░®ļÉśņ¢┤ ņ׳ņŚłņ£╝ļ®░(Fig. 1C), ņØ╝ļČĆ Ļ░£ņ▓┤ņØś ļ░®Ļ┤æ ļé┤ņŚÉņä£ļÅä Ļ┤Ćņ░░ĒĢĀ ņłś ņ׳ņŚłļŗż.
ĒÖĢņ¦äļÉ£ 153ļ¦łļ”¼ ļÅ╝ņ¦ĆņŚÉņä£ ņåīņן ļé┤ ņłśņ¢æņä▒ ļé┤ņÜ®ļ¼╝ņØś ņĀĆļźś(98.0%)Ļ░Ć ņ£ä ļé┤ ņØæņ£ĀņØś ņĪ┤ņ×¼(62.7%) ļ░Å ņŗĀņןņØś ņÜöņé░ņŚ╝ ņ╣©ņ░®(60.1%)ņŚÉ ļ╣äĒĢśņŚ¼ ļåÆņØĆ ļ╣łļÅäļĪ£ Ļ┤Ćņ░░ļÉśņŚłļŗż(Table 2). ļÅ╝ņ¦ĆņØś ņŚ░ļĀ╣ļ│äļĪ£ļŖö Ēü░ ņ░©ņØ┤ļź╝ ļ│┤ņØ┤ņ¦ĆļŖö ņĢŖņĢśņ¦Ćļ¦ī, ņŗĀņןņØś ņÜöņé░ņŚ╝ ņ╣©ņ░®ņØĆ 8ņØ╝ļĀ╣ ņØ┤ņāüņØś ļÅ╝ņ¦ĆņŚÉņä£ ĒśäņĀĆĒĢśĻ▓ī ļé«ņĢśļŗż.
ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀü Ļ▓Ćņé¼ Ļ▓░Ļ│╝
ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀüņ£╝ļĪ£ ņåīņןņØś ņĀÉļ¦ēņĖĄņŚÉņä£ Ļ░Ćņן ĒŖ╣ņ¦ĢņĀüņØĖ ļ│ĆĒÖöņāüņØ┤ Ļ┤Ćņ░░ļÉśņŚłņ£╝ļ®░, ņĀÉļ¦ēĒĢśņ¦ü, ĻĘ╝ņ£ĪņĖĄ ļ░Å ņןļ¦ēņŚÉņä£ļŖö ĒŖ╣ļ│äĒĢ£ ļ│æļ│ĆņØä Ļ┤Ćņ░░ĒĢĀ ņłś ņŚåņŚłļŗż. PEDļĪ£ ĒÖĢņ¦äļÉ£ ļÅ╝ņ¦ĆņØś ņåīņןņŚÉņä£ Ļ░Ćņן ļæÉļō£ļ¤¼ņ¦ä ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀü ļ│æļ│ĆņØĆ ņ£Ąļ¬©ņØś ĒśäņĀĆĒĢ£ ņ£äņČĢĻ│╝ ņ£ĄĒĢ®ņØ┤ņŚłļŗż. Ļ│ĄņןĻ│╝ ĒÜīņןņŚÉ ļīĆĒĢśņŚ¼ VH:CD ļ╣äņ£©ņØä ļ╣äĻĄÉĒĢ£ Ļ▓░Ļ│╝ Ļ│Ąņן(1.4:1)ņØ┤ ĒÜīņן(1.5:1)ņŚÉ ļ╣äĒĢśņŚ¼ ļŗżņåī ļé«Ļ▓ī ļéśĒāĆļé¼ņ£╝ļ®░, ļŗżļźĖ ņŚ░ļĀ╣ņŚÉ ļ╣äĒĢśņŚ¼ 4-7ņØ╝ļĀ╣ņØś ļÅ╝ņ¦ĆļōżņØ┤ Ļ│ĄņןĻ│╝ ĒÜīņן ļ¬©ļæÉņŚÉņä£ 1.3:1ļĪ£ Ļ░Ćņן ļé«ņĢśļŗż(Table 3).
ļ░öņØ┤ļ¤¼ņŖż Ļ░ÉņŚ╝ņŚÉ ļö░ļźĖ ņåīņן ņ£Ąļ¬©ņØś ņ£äņČĢļ┐Éļ¦ī ņĢäļŗłļØ╝, ņåÉņāü ļ░øņØĆ ņ£Ąļ¬©Ļ░Ć ņä£ļĪ£ ņ£ĄĒĢ®ĒĢśļŖö ļ│æļ│ĆļÅä ņ×ÉņŻ╝ Ļ┤Ćņ░░ļÉśņŚłļŗż. ļČĆņ£äļ│äļĪ£ļŖö Ļ│Ąņן(72.6%, 90ļ¦łļ”¼)ļ│┤ļŗżļŖö ĒÜīņן(78.2%, 97ļ¦łļ”¼)ņŚÉņä£ ņ£Ąļ¬©ņØś ņ£ĄĒĢ®ņØ┤ ļŹöņÜ▒ ļ╣łļ▓łĒĢśĻ▓ī Ļ┤Ćņ░░ļÉśņŚłļŗż(Table 4). ļśÉĒĢ£ ņŗ¼ĒĢ£ ļ│æļ│Ć ļō▒ĻĖēņØĖ G3Ļ░Ć Ļ│Ąņן(50.0%, 62ļ¦łļ”¼)Ļ│╝ ĒÜīņן(54.0%, 67ļ¦łļ”¼)ņŚÉņä£ ļ¬©ļæÉ ļåÆĻ▓ī Ļ┤Ćņ░░ļÉśņŚłļŗż.
ļśÉĒĢ£ ņ£Ąļ¬©ņāüĒö╝ņäĖĒżņØś ļ»ĖņäĖņ£Ąļ¬© ņåīņŗż, ņ×ģļ░®ĒÖö(Fig. 2A), ĒÄĖĒÅēĒÖö(Fig. 2B), ņāüĒö╝ņäĖĒżņØś ņäĖĒżņ¦łņŚÉ ĒĢśļéś ļśÉļŖö ņŚ¼ļ¤¼ Ļ░£ņØś Ēł¼ļ¬ģĒĢ£ Ļ│ĄĒż ĒśĢņä▒(Fig. 2C) ļ░Å ņāüĒö╝ņäĖĒż ĒāłļØĮ(Fig. 2D)ņØ┤ Ļ┤Ćņ░░ļÉśņŚłļŗż. ņØ╝ļČĆ Ļ░£ņ▓┤ņŚÉņä£ļŖö ņøĆņØś ņןņäĀņØ┤ ĒśĖņŚ╝ņä▒ņ£╝ļĪ£ ļåŹņŚ╝ļÉśņ¢┤ ņ׳Ļ│Ā, ņłĀņ×öņäĖĒżĻ░Ć Ļ▒░ņØś ņåīņŗżļÉ£ ņ×¼ņāØņØś ĒØöņĀüņØ┤ ņĪ┤ņ×¼ĒĢśĻ│Ā ņ׳ņŚłļŗż. ĻĘĖļ¤¼ļéś Ļ│Āņ£ĀņĖĄņŚÉņä£ņØś ņŚ╝ņ”Ø ļ░śņØæņØĆ Ļ▒░ņØś ņŚåĻ▒░ļéś ļ¦żņÜ░ ļ»ĖņĢĮĒĢśņśĆļŗż. ņŗĀņן ņłśņ¦łļČĆņØś ņøÉņ£äĻ│ĪņäĖļć©Ļ┤Ć ļśÉļŖö ņ¦æĒĢ®Ļ┤Ć ļé┤Ļ░ĢņŚÉ ĒśĖņé░ņä▒ņØś Ļ│╝ļ”Įņāü ļ¼╝ņ¦łņØĖ ņÜöņé░ņØ┤ ņ×ÉņŻ╝ Ļ┤Ćņ░░ļÉśņŚłļŗż. Ļ│Ąņן 126ļ¦łļ”¼ ļ░Å ĒÜīņן 120ļ¦łļ”¼ņŚÉņä£ Ļ┤Ćņ░░ļÉ£ ņ£Ąļ¬©ņāüĒö╝ņäĖĒżņØś ĒŖ╣ņ¦ĢņĀü ļ│æļ│ĆļōżņØä ļ╣äĻĄÉĒĢśņśĆļŗż(Table 5). ņØ┤ ņżæ ņäĖĒżņ¦łņŚÉ Ļ│ĄĒż ĒśĢņä▒ ļ│æļ│ĆņØ┤ Ļ░Ćņן ļåÆņØĆ ļ╣äņ£©ļĪ£ Ļ┤Ćņ░░ļÉśņŚłĻ│Ā ĒÄĖĒÅēĒÖö ļ░Å ņāüĒö╝ ĒāłļØĮ ņł£ņ£╝ļĪ£ ļéśĒāĆļé¼ļŗż. ĒŖ╣Ē׳ ņäĖĒżņ¦łņØś Ļ│ĄĒżĒÖöļŖö G3 ļō▒ĻĖē ļ╣äņ£©ņØ┤ Ļ░Ćņן ļåÆņĢśĻ│Ā, Ļ│Ąņן(75.4%, 95ļ¦łļ”¼)ļ│┤ļŗżļŖö ĒÜīņן(81.7%, 98ļ¦łļ”¼)ņŚÉņä£ ļŹöņÜ▒ ļ╣łļ▓łĒĢśĻ▓ī ĒÖĢņØĖļÉśņŚłļŗż. ņāüĒö╝ņäĖĒżņØś ĒāłļØĮņØĆ Ļ│ĄņןĻ│╝ ĒÜīņן ļ¬©ļæÉņŚÉņä£ G1 ļśÉļŖö G3 ļ╣äņ£©ņØ┤ ļåÆņØĆ ļ░śļ®┤, ĒÄĖĒÅēĒÖö ļ│æļ│ĆņØĆ Ļ│ĄņןņŚÉņä£ņØś G3 ļ╣äņ£©ņØ┤ ĒÜīņןņŚÉ ļ╣äĒĢśņŚ¼ ļåÆĻ▓ī Ļ┤Ćņ░░ļÉśņŚłļŗż. ļÅ╝ņ¦ĆņØś ņŚ░ļĀ╣ļ│ä ļ╣äĻĄÉĒĢśļ®┤, Ļ│ĄņןņŚÉņä£ļŖö ļÅ╝ņ¦ĆņØś ņØ╝ļĀ╣ņŚÉ ņāüĻ┤ĆņŚåņØ┤ ņāüĒö╝ņäĖĒżņØś Ļ│ĄĒżĒÖöĻ░Ć Ļ░Ćņן ļ¦ÄņØ┤ Ļ┤Ćņ░░ļÉśņŚłņ£╝ļ®░ ĒÄĖĒÅēĒÖö ļ░Å ņāüĒö╝ņäĖĒżĒāłļØĮ ņł£ņ£╝ļĪ£ ļéśĒāĆļé¼ļŗż. Ļ│ĄĒżĒÖöņÖĆ ĒÄĖĒÅēĒÖö ļ│æļ│ĆņØĆ G3 ļō▒ĻĖēņØ┤ ļ¦ÄņØĆ ļ╣łļÅäļź╝ ņ░©ņ¦ĆĒĢśļŖö ļ░śļ®┤, ņāüĒö╝ņäĖĒżņØś ĒāłļØĮ ļ│æļ│ĆņØĆ ļō▒ĻĖēļ│äļĪ£ ļ╣äĻĄÉņĀü ļŗżņ¢æĒĢśĻ▓ī Ļ┤Ćņ░░ļÉśņŚłļŗż. ĒÜīņןņØś Ļ▓ĮņÜ░ 4ņØ╝ļĀ╣ ļ»Ėļ¦ī ļ░Å 4-7ņØ╝ļĀ╣ņØś ļÅ╝ņ¦ĆņŚÉņä£ļŖö ņāüĒö╝ņäĖĒżņØś Ļ│ĄĒżĒÖöĻ░Ć Ļ░Ćņן ļ¦ÄņØ┤ Ļ┤Ćņ░░ļÉśļŖöļŹ░ ļ╣äĒĢśņŚ¼, 8ņØ╝ļĀ╣ ņØ┤ņāüņŚÉņä£ļŖö ļÜ£ļĀĘĒĢ£ ņ░©ņØ┤ļź╝ ļéśĒāĆļé┤Ļ│Ā ņ׳ņ¦Ć ņĢŖņĢśļŗż. ļśÉĒĢ£ Ļ│ĄĒżĒÖö ļ│æļ│ĆņØĆ 4ņØ╝ļĀ╣ ļ»Ėļ¦ī ļ░Å 4-7ņØ╝ļĀ╣ņØś ļÅ╝ņ¦ĆņŚÉņä£ G3ļō▒ĻĖē ļ░£ĒśäņØ┤ ļåÆĻ▓ī ļéśĒāĆļé¼ļŗż.
ļ®┤ņŚŁņĪ░ņ¦üĒÖöĒĢÖ Ļ▓Ćņé¼ Ļ▓░Ļ│╝
ņåīņןņŚÉņä£ PEDV ĒĢŁņøÉņØĆ ļīĆļČĆļČä ņ£äņČĢļÉ£ ņ£Ąļ¬© ļśÉļŖö ņĀÉļ¦ēņØä Ēö╝ļ│ĄĒĢśĻ│Ā ņ׳ļŖö Ļ│ĄĒżĒÖö ļ░Å ņ×ģļ░®ĒÖöĻ░Ć ņ¦äĒ¢ēļÉ£ ņāüĒö╝ņäĖĒżņØś ņäĖĒżņ¦ł ļśÉļŖö ņåöļ│ĆņŚ░ņŚÉņä£ ļ░£ĒśäļÉśĻ│Ā ņ׳ņŚłņ£╝ļ®░(Fig. 3A and B), ņØ╝ļČĆ ĒāłļØĮ ņāüĒö╝ ļśÉļŖö ņøĆ ņāüĒö╝ņäĖĒżņŚÉņä£ļÅä ĒĢŁņøÉņØä Ļ┤Ćņ░░ĒĢĀ ņłś ņ׳ņŚłļŗż. ņĪ░ņ¦üĒĢÖņĀüņ£╝ļĪ£ ĒĢŁņøÉņØĆ EG1 ļśÉļŖö EG2ļĪ£ ļČäĒżĒĢśĻ▒░ļéś ņŗ¼ĒĢ£ Ļ▓ĮņÜ░ EG3ņØś ļČäĒżļź╝ ļ│┤ņØ┤ĻĖ░ļÅä ĒĢśņśĆļŗż(Table 6). ĻĘĖļ¤¼ļéś ņĀÉļ¦ēĻ│Āņ£ĀņĖĄ, ņĀÉļ¦ēĒĢśņ¦ü ļ░Å ĻĖ░ĒāĆ ĻĘ╝ņ£ĪņĖĄņŚÉņä£ļŖö ļ░öņØ┤ļ¤¼ņŖż ĒĢŁņøÉņØä Ļ▓ĆņČ£ĒĢĀ ņłś ņŚåņŚłļŗż. ĒŖ╣Ē׳ PEDVņØś Ēæ£ņĀüņןĻĖ░Ļ░Ć ņĢäļŗī Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŖö Ļ▓░ņןņØś ņĀÉļ¦ēņāüĒö╝ņäĖĒżņŚÉņä£ļÅä ļ░öņØ┤ļ¤¼ņŖż ĒĢŁņøÉņØ┤ Ļ▓ĆņČ£ļÉśĻĖ░ļÅä ĒĢśņśĆņ£╝ļ®░(Fig. 3C), ņåīņןĻ│╝ ļŗ¼ļ”¼ ĻĄŁņåīņĀüņØĖ ļČäĒżļź╝ ļ│┤ņØ┤Ļ│Ā ņ׳ņŚłļŗż.
ļ░öņØ┤ļ¤¼ņŖż ĒĢŁņøÉ ļ░£ĒśäņØĆ ĒÜīņן(85.8%, 109ļ¦łļ”¼)ņØ┤ Ļ│Ąņן(82.2%, 111ļ¦łļ”¼)ņŚÉ ļ╣äĒĢśņŚ¼ ļåÆĻ▓ī ļéśĒāĆļé¼ļŗż(Table 6). ņĀäņ▓┤ņĀüņ£╝ļĪ£ ņåīņן ņĪ░ņ¦üņŚÉņä£ļŖö EG3 ļō▒ĻĖēņØś ĒĢŁņøÉ ļ░£Ēśä Ļ░£ņ▓┤Ļ░Ć 80% ņØ┤ņāüņØä ņĀÉņ£ĀĒĢśĻ│Ā ņ׳ņŚłļŗż. Ļ▓░ņןņØś Ļ▓ĮņÜ░ Ļ▓Ćņé¼ĒĢ£ 133ļ¦łļ”¼ Ļ░£ņ▓┤ ņżæ 19.5% (26ļ¦łļ”¼)ņŚÉņä£ ĒĢŁņøÉņØ┤ Ļ▓ĆņČ£ļÉśņŚłļŗż. Ļ░ü ņןĻĖ░ļ│äļĪ£ ĒÅēĻĘĀ ĒĢŁņøÉ ļ░£Ēśä ļō▒ĻĖēņØä ļ╣äĻĄÉĒĢ£ Ļ▓░Ļ│╝, Ļ│ĄņןĻ│╝ ĒÜīņןņØ┤ Ļ▓░ņןņŚÉ ļ╣äĒĢ┤ņä£ ņ£ĀņØśņä▒ ņ׳Ļ▓ī ļåÆņØĆ ņłśņżĆņ£╝ļĪ£ ļ░£ĒśäļÉśņŚłļŗż(p < 0.05). ĒĢŁņøÉņØś ļ░£ĒśäņØä ļÅ╝ņ¦ĆņØś ņŚ░ļĀ╣ļ│äļĪ£ ļ╣äĻĄÉĒĢ£ Ļ▓░Ļ│╝(Table 5), 4-7ņØ╝ļĀ╣ņØś ļÅ╝ņ¦Ć Ļ│Ąņן(90.0%, 36ļ¦łļ”¼)Ļ│╝ ĒÜīņן(92.1%, 35ļ¦łļ”¼)ņŚÉņä£ Ļ░Ćņן ļ¦ÄņØ┤ Ļ▓ĆņČ£ļÉśņŚłļŗż. ļ░śļ®┤ 8ņØ╝ļĀ╣ ņØ┤ņāü ļÅ╝ņ¦ĆņØś Ļ│Ąņן(55.6%, 5ļ¦łļ”¼)Ļ│╝ ĒÜīņן(66.7%, 6ļ¦łļ”¼) ļ¬©ļæÉņŚÉņä£ ĒśäņĀĆĒĢśĻ▓ī Ļ░ÉņåīĒĢśļŖö Ļ▓ĮĒ¢źņØä ļ│┤ņØ┤Ļ│Ā ņ׳ņŚłļŗż. ļśÉĒĢ£ Ļ│Ąņן ļ░Å ĒÜīņןņØś PEDV ĒÅēĻĘĀ ĒĢŁņøÉ ļ░£Ēśä ļō▒ĻĖēņØä ņŚ░ļĀ╣ļ│äļĪ£ ļ╣äĻĄÉĒĢ£ Ļ▓░Ļ│╝ņŚÉņä£ļÅä 8ņØ╝ļĀ╣ ļ»Ėļ¦īņØś ļÅ╝ņ¦ĆĻ░Ć 8ņØ╝ļĀ╣ ņØ┤ņāü ļÅ╝ņ¦ĆņØś ļ░£Ēśä ļō▒ĻĖēļ│┤ļŗż ņ£ĀņØśņä▒ ņ׳Ļ▓ī ļåÆĻ▓ī ļéśĒāĆļé¼ļŗż(p < 0.05).
Ļ▓░ņןņØś Ļ▓ĮņÜ░ 4ņØ╝ļĀ╣ ļ»Ėļ¦ī 19ļ¦łļ”¼, 4-7ņØ╝ļĀ╣ 6ļ¦łļ”¼ ļ░Å 8ņØ╝ļĀ╣ ņØ┤ņāü 1ļ¦łļ”¼ ļō▒ ņ┤Ø 26ļ¦łļ”¼ņØś ļÅ╝ņ¦ĆņŚÉņä£ PEDV ĒĢŁņøÉņØ┤ Ļ▓ĆņČ£ļÉśņŚłļŗż.
ļ│æņøÉņ▓┤ ļ░Å ņ£ĀņĀäĒĢÖņĀü Ļ▓Ćņé¼ Ļ▓░Ļ│╝
ļÅ╝ņ¦Ć ļČäļ│ĆņŚÉ ļīĆĒĢ£ RT-PCR Ļ▓Ćņé¼ Ļ▓░Ļ│╝, PEDV ļ░Å TGEV ņ£ĀņĀäņ×É ņ”ØĒÅŁņé░ļ¼╝ņØ┤ Ļ░üĻ░ü 525 bp ļ░Å 755 bpņŚÉņä£ Ļ▓ĆņČ£ļÉśņŚłļŗż(Fig. 3D). TGEVļŖö ņĀä Ļ░£ņ▓┤ ņØīņä▒ņØ┤ņŚłņ£╝ļ®░, PEDVļŖö 151ļ¦łļ”¼(98.7%)ņŚÉņä£ ņ¢æņä▒ ļ░śņØæņØä ļ│┤ņśĆņ£╝ļéś ļéśļ©Ėņ¦Ć 2ļæÉ(3 ļ░Å 4ņØ╝ļĀ╣)ļŖö ņØīņä▒ņ£╝ļĪ£ ĒÖĢņØĖļÉśņŚłļŗż.
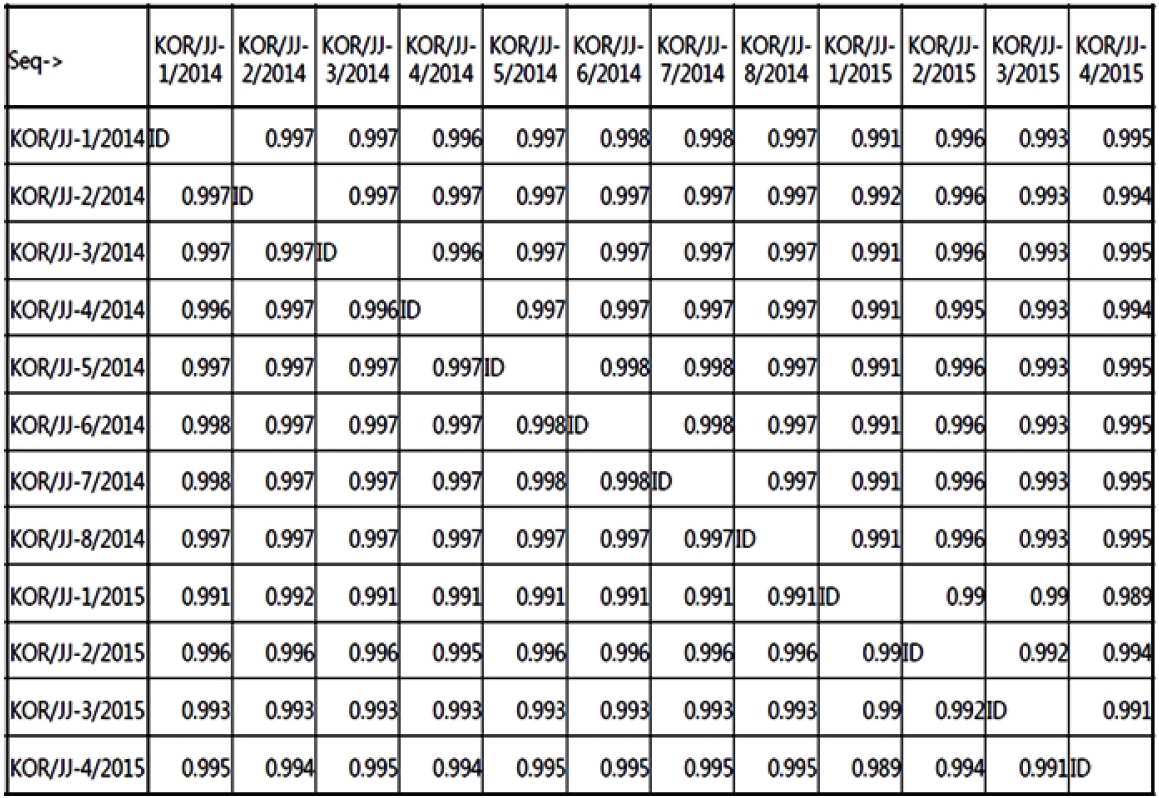
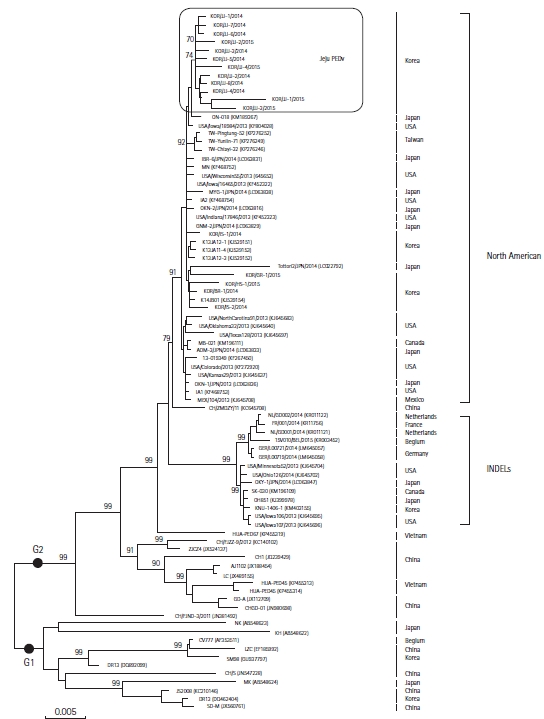
ņĀ£ņŻ╝ļÅäņŚÉņä£ Ļ▓ĆņČ£ļÉ£ 12ņŻ╝ņØś PEDV S ņ£ĀņĀäņ×Éļź╝ ļČäņäØĒĢ£ Ļ▓░Ļ│╝ Ļ░ü ļČäļ”¼ņŻ╝Ļ░äņŚÉ 98.9% ņØ┤ņāüņØś ņāüļÅÖņä▒ņØä ļ│┤ņØ┤Ļ│Ā ņ׳ņŚłļŗż(Fig. 4). ļśÉĒĢ£ Ļ│äĒåĄĒĢÖņĀü ĒŖĖļ”¼ļź╝ ņ×æņä▒ĒĢ£ Ļ▓░Ļ│╝, ļČäļ”¼ļÉ£ PEDVļŖö genogroup 2 (G2)ņŚÉ ņåŹĒĢśņŚ¼ ņ׳ņØīņØ┤ ņ×ģņ”ØļÉśņŚłļŗż(Fig. 5). ņĄ£ĻĘ╝ ļ»ĖĻĄŁņŚÉņä£ļŖö ļ│æņøÉņä▒ņØ┤ Ļ▓Įļ»ĖĒĢ£ insertions and delections in the spike gene (INDELs) ĻĘĖļŻ╣ PEDVņÖĆ ļ│æņøÉņä▒ņØ┤ Ļ░ĢĒĢ£ North American ĻĘĖļŻ╣ PEDV 2ņóģņØ┤ ļČäļ”¼ļÉśņŚłņ£╝ļ®░[11], ņØ┤ļ▓ł ņŚ░ĻĄ¼ņŚÉņä£ ļČäņäØĒĢ£ 12ņŻ╝ņØś PEDVļŖö ļ¬©ļæÉ North American ĻĘĖļŻ╣ņŚÉ ņåŹĒĢśĻ│Ā ņ׳ņØīņØä ĒÖĢņØĖĒĢĀ ņłś ņ׳ņŚłļŗż.
Ļ│Āņ░░
ļÜ£ļĀĘĒĢ£ ļÅÖņĀłĻĖ░ņØś Ļ│äņĀł ļ░£ņāØņØä ļ│┤ņØ┤ļŖö TGEņÖĆļŖö ļŗ¼ļ”¼ Ļ│╝Ļ▒░ ĻĄŁļé┤ņŚÉņä£ ņ£ĀĒ¢ēĒĢśļŹś PEDļŖö Ļ▒░ņØś ņŚ░ņżæ ļ░£ņāØĒĢśņśĆņ¦Ćļ¦ī, ļīĆņ▓┤ļĪ£ ļÅÖņĀłĻĖ░ņŚÉ ĒĢ┤ļŗ╣ĒĢśļŖö ĻĖ░Ļ░äņØ┤ 11ņøöņŚÉņä£ 3ņøöĻ╣īņ¦Ć ņ¦æņżæĒĢśņŚ¼ ļ░£ņāØĒĢśļŖö Ļ▓ĮĒ¢źņØä ļ│┤ņØ┤Ļ│Ā ņ׳ņŚłļŗż[5]. ņØ┤ļ▓ł ņŚ░ĻĄ¼ļź╝ ĒåĄĒĢ┤ ņĀ£ņŻ╝ļÅä ļé┤ņŚÉņä£ PEDļŖö ņ×¼ļ░£ņāØ ņŗ£ņĀÉņŚÉ ĒÅŁļ░£ņĀüņØĖ ļ░£ņāØņØ┤ ņ׳ņØĆ Ēøä Ļ▒░ņØś Ļ│äņĀłņŚÉ Ļ┤ĆĻ│äņŚåņØ┤ ņŚ░ņżæ ļ░£ņāØĒĢśļŗżĻ░Ć ļÅÖņĀłĻĖ░ņŚÉ ļōżņ¢┤ņä£ļ®┤ņä£ ņ”ØĻ░ĆĒĢśļŖö Ļ▓ĮĒ¢źņØä ļ│┤ņØ┤Ļ│Ā ņ׳ņŚłļŗż. ļśÉĒĢ£ PED ļ░£ņāØ 52ļåŹĻ░Ć ņżæ 17Ļ░£(32.7%) ļåŹĻ░ĆņŚÉņä£ 2ĒÜīņŚÉņä£ ļ¦ÄĻ▓īļŖö 4ĒÜīĻ╣īņ¦Ć ņ¦ĆņåŹ ļ░£ņāØĒĢśļŖö ņ¢æņāüņØä ļéśĒāĆļé┤Ļ│Ā ņ׳ņŚłļŗż. ļŹöņÜ▒ņØ┤ ņ¢æļÅł ļåŹĻ░ĆĻ░Ć ļ░Ćņ¦æļÉśņ¢┤ ņ׳ļŖö ņĀ£ņŻ╝ļÅä ņä£ļČĆņ¦ĆņŚŁ(ĒĢ£ļ”╝ņØŹ, ļīĆņĀĢņØŹ ļ░Å ĒĢ£Ļ▓Įļ®┤)ņŚÉņä£ ņ¦æņżæņĀüņ£╝ļĪ£ ļ░£ņāØĒĢśĻ│Ā ņ׳ņŚłļŗż. ņØ┤ļ¤¼ĒĢ£ ņĀÉņØĆ ļåŹņן Ļ░äņØś ņ¦łļ│æ ņĀäĒīīļź╝ ņśłļ░®ĒĢśĻĖ░ ņ£äĒĢ£ ņ░©ļŗ©ļ░®ņŚŁņØ┤ ņĀ£ļīĆļĪ£ ņłśĒ¢ēļÉśĻ│Ā ņ׳ņ¦Ć ņĢŖņØä ļ┐Éļ¦ī ņĢäļŗłļØ╝, ļåŹņןņŚÉņä£ņØś ļČĆņŗżĒĢ£ ņśłļ░®ņĀæņóģ, ņ¦łļ│æ ļ░£ņāØ ļåŹņןņØś ņØĖĻ│Ą ļ®┤ņŚŁ ņŗżņŗ£ ļśÉļŖö ļČĆņĀüņĀłĒĢ£ ĒÖśļÅł Ļ┤Ćļ”¼ ļō▒ņØ┤ ņ¦ĆņåŹņĀü PED ļ░£ņāØņØś ņøÉņØĖņ£╝ļĪ£ ņČöņĀĢļÉśĻ│Ā ņ׳ļŗż. Ļ│╝Ļ▒░ ĻĄŁļé┤ ņ¢æļÅłņןņØä ļīĆņāüņ£╝ļĪ£ ņŗżņŗ£ĒĢ£ PED ņŚŁĒĢÖ ņĪ░ņé¼ Ļ▓░Ļ│╝ņŚÉ ļö░ļź┤ļ®┤, PEDVņØś ļåŹņן ļé┤ ņł£ĒÖś Ļ░ÉņŚ╝ņØś ņøÉņØĖņ£╝ļĪ£ ļČäļ¦īņé¼ ĒÖśļÅłņŚÉ ļīĆĒĢ£ ļČĆņĀüņĀłĒĢ£ ņĪ░ņ╣ś, ļČĆņĀüņĀłĒĢ£ ņśłļ░®ņĀæņóģņŚÉ ļö░ļźĖ ļ¬©ļÅłņØś ļ®┤ņŚŁ ņłśņżĆ ļČłĻĘĀĒśĢ, ņ£Āņ”ÖļČłļ¤ē ļ¬©ļÅłņŚÉņä£ ņ×ÉļÅłņ£╝ļĪ£ ņ┤łņ£ĀĒĢŁņ▓┤ ņĀäļŗ¼ ņŗżĒī©, ļ¬©ļÅłņłś Ļ│╝ļŗżņŚÉ ļö░ļźĖ ļČäļ¦īņé¼ Ļ│ĄĻ░ä ļČĆņĪ▒ ļō▒ņ£╝ļĪ£ ļČäņäØļÉ£ ļ░ö ņ׳ļŗż[12].
PEDVļŖö ļÅ╝ņ¦ĆņØś ņåīņן ĒŖ╣Ē׳, Ļ│Ąņן ļ░Å ĒÜīņןņØś ņ£Ąļ¬©ņāüĒö╝ņäĖĒżņŚÉ Ļ░ÉņŚ╝ļÉśņ¢┤ ņ£Ąļ¬©ņØś ņ£äņČĢņØä ņ┤łļלĒĢśņŚ¼ ĒØĪņłśļČłļ¤ēņä▒ ņäżņé¼ļź╝ ņ£Āļ░£ĒĢ£ļŗż. PEDV Ļ░ÉņŚ╝ ņŗ£ ņ£ĪņĢłņĀüņ£╝ļĪ£ ņåīņןņØĆ ĒīĮņ£żļÉśĻ│Ā ĒÖ®ņāēņØś ņłśņ¢æņä▒ ļé┤ņÜ®ļ¼╝ņØä ĒĢ©ņ£ĀĒĢśĻ│Ā ņ׳ņ£╝ļ®░, ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀüņ£╝ļĪ£ļŖö VH:CD ļ╣äņ£©ņØ┤ ņĀĢņāüļ▓öņ£äņØĖ 7-8:1ņŚÉņä£ 3:1Ļ╣īņ¦Ć Ļ░ÉņåīĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[2]. ĻĄŁļé┤ņŚÉņä£ļŖö PED Ļ░ÉņŚ╝ņØ┤ ņ▓śņØīņ£╝ļĪ£ ĒÖĢņØĖļÉ£ 1992-1993ļģä ņé¼ņØ┤ ņ┤Ø 71Ļ▒┤ņØä Ļ▓Ćņé¼ĒĢ£ Ļ▓░Ļ│╝ Ļ│Ąņן ļ░Å ĒÜīņןņŚÉņä£ VH:CD ļ╣äņ£©ņØ┤ ĒÅēĻĘĀ 3:1-2:1 ņĀĢļÅäļĪ£ Ļ░ÉņåīĒĢśņśĆņ£╝ļ®░, ņŗ¼ĒĢ£ ņśłņŚÉņä£ļŖö 1:1ņŚÉņä£ 1:2Ļ╣īņ¦Ć ĒśäņĀĆĒĢśĻ▓ī ņ£äņČĢļÉśĻĖ░ļÅä ĒĢśņśĆļŗż[5]. ņĄ£ĻĘ╝ 2013ļģä ļ»ĖĻĄŁņŚÉņä£ļŖö PEDļĪ£ ĒÖĢņ¦äļÉ£ ņ×ÉļÅłņØś ņןļé┤ņÜ®ļ¼╝ņŚÉņä£ ļČäļ”¼ļÉ£ PEDVļź╝ 10-35ņØ╝ļĀ╣ ļ¼┤ĻĘĀļÅ╝ņ¦ĆņŚÉ Ļ▓ĮĻĄ¼Ļ░ÉņŚ╝ņŗ£Ēé© Ļ▓░Ļ│╝ ņĀæņóģ 24-48ņŗ£Ļ░ä Ēøä ņŗ¼ĒĢ£ ņäżņé¼ ļ░Å ĻĄ¼ĒåĀ ņ”ØņāüņØä ļ│┤ņśĆņ£╝ļ®░, ņåīņןņŚÉņä£ VH:CD ļ╣äņ£©ņØ┤ Ļ░£ņ▓┤ļ│äļĪ£ 1.2-3.4:1 ņĀĢļÅäļĪ£ ņŗ¼ĒĢ£ ņ£Ąļ¬©ņØś ņ£äņČĢņØä ļ│┤ņśĆļŗżĻ│Ā ĒĢśņśĆļŗż[6]. ņØ┤ļ▓ł ņŚ░ĻĄ¼ņŚÉņä£ ņĀ£ņŻ╝ļÅä ļÅ╝ņ¦ĆņØś Ļ▓ĮņÜ░ Ļ│ĄņןĻ│╝ ĒÜīņןņØś ĒÅēĻĘĀ VH:CD ļ╣äņ£©ņØ┤ Ļ░üĻ░ü 1.4:1 ļ░Å 1.5:1ļĪ£ ļéśĒāĆļé¼ņ£╝ļ®░, ĒŖ╣Ē׳ 4-7ņØ╝ļĀ╣ ņ×ÉļÅłņØś Ļ│ĄņןĻ│╝ ĒÜīņן ļ¬©ļæÉņŚÉņä£ 1.3:1ļĪ£ Ļ░Ćņן ļé«Ļ▓ī ļéśĒāĆļé¼ļŗż. ļśÉĒĢ£ ņ£äņČĢļÉ£ ņ£Ąļ¬©ņØś ņ£ĄĒĢ® ņåīĻ▓¼ļÅä ņŗ¼ĒĢ£ ļō▒ĻĖēņØĖ G3ņØś ļ╣äņ£©ņØ┤ 50% ņØ┤ņāüņØś ļÅ╝ņ¦Ć Ļ│Ąņן ļ░Å ĒÜīņןņŚÉņä£ Ļ┤Ćņ░░ļÉśĻ│Ā ņ׳ņŚłļŗż. ļö░ļØ╝ņä£ Ļ│╝Ļ▒░ ĻĄŁļé┤ņŚÉņä£ ļ░£ņāØĒĢ£ PEDVņŚÉ ļ╣äĒĢśņŚ¼ ņĄ£ĻĘ╝ ņĀ£ņŻ╝ļÅäņŚÉņä£ ļ░£ņāØĒĢśĻ│Ā ņ׳ļŖö PEDVņØś ļ│æņøÉņä▒ņØ┤ ņóĆ ļŹö Ļ░ĢĒĢĀ Ļ▓āņ×äņØä Ļ░äņĀæņĀüņ£╝ļĪ£ ņŗ£ņé¼ĒĢśĻ│Ā ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉ£ļŗż.
PEDVņŚÉ Ļ░ÉņŚ╝ļÉ£ ļÅ╝ņ¦ĆņØś ņåīņןņŚÉņä£ļŖö ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀüņ£╝ļĪ£ ņן ņ£Ąļ¬©ņØś ņ£äņČĢĻ│╝ ĒĢ©Ļ╗ś ņāüĒö╝ņäĖĒżņØś ĒāłļØĮ ļ░Å ņäĖĒżņ¦ł ļé┤ Ļ│ĄĒż ĒśĢņä▒ ņåīĻ▓¼ņØ┤ ĒŖ╣ņ¦ĢņĀüņ£╝ļĪ£ ļéśĒāĆļé£ļŗż[2,6]. Ļ│╝Ļ▒░ ĻĄŁļé┤ ļ░£ņāØĒĢ£ PED Ļ░ÉņŚ╝ ļÅ╝ņ¦ĆņŚÉņä£ ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀü ņåīĻ▓¼ņØĆ ņŗŁņØ┤ņ¦Ćņןļ│┤ļŗżļŖö Ļ│ĄņןĻ│╝ ĒÜīņןņŚÉņä£ ņŻ╝ļĪ£ ļ│æļ│ĆņØ┤ Ļ┤Ćņ░░ļÉśņŚłļŗż. ņ£Ąļ¬©ņāüĒö╝ņäĖĒżņØś ņäĖĒżņ¦łņŚÉ ļ¼┤ņłśĒ׳ ļ¦ÄņØĆ ņ×æņØĆ Ļ│ĄĒżĻ░Ć ĒśĢņä▒ļÉśņ¢┤ ņ׳Ļ│Ā, ņøÉņŻ╝ņāüĒö╝ņäĖĒżņØś ļåÆņØ┤Ļ░Ć ļé«ņĢäņĀĖ ņ×ģļ░®ĒÖöļÉśĻ▒░ļéś ņŗ¼ĒĢ£ Ļ▓ĮņÜ░ ĒÄĖĒÅēĒĢśĻ▓ī ļÉśņŚłņ£╝ļ®░, ļĢīļĪ£ ņāüĒö╝ņäĖĒżĻ░Ć ļ░Ģļ”¼ļÉśņ¢┤ ņĀÉļ¦ēĻ│Āņ£ĀņĖĄņØ┤ ļģĖņČ£ļÉśņ¢┤ ņ׳ĻĖ░ļÅä ĒĢśņśĆļŗż[5,13]. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀü ņåīĻ▓¼ņØś ļ░£ņāØ ļ╣łļÅäļź╝ ņé┤ĒÄ┤ļ│┤ņĢśņØä ļĢī, ņäĖĒżņ¦ł ļé┤ Ļ│ĄĒż ĒśĢņä▒, ņāüĒö╝ņäĖĒżņØś ņ×ģļ░®ĒÖö ļśÉļŖö ĒÄĖĒÅēĒÖö ļ░Å ĒāłļØĮņØś ņł£ņ£╝ļĪ£ ļéśĒāĆļéśĻ│Ā ņ׳ņŚłļŗż. ļśÉĒĢ£ ņŗ¼ĒĢ£ Ļ░£ņ▓┤ņŚÉņä£ļŖö ļ»ĖņäĖņ£Ąļ¬©ņØś ņåīņŗż ļśÉĒĢ£ Ļ┤Ćņ░░ļÉśņŚłļŗż. ĻĘĖļ¤¼ļ»ĆļĪ£ PEDV Ļ░ÉņŚ╝ņŚÉ ļö░ļźĖ ņ£Ąļ¬©ņØś ņ£äņČĢ, ņāüĒö╝ņäĖĒżņØś Ļ│ĄĒżĒÖö ļ░Å ņØ┤ņŚÉ ļö░ļź┤ļŖö ņØ╝ļĀ©ņØś ļ│ĆĒÖöļĪ£ ņØĖĒĢśņŚ¼ ņåīņןņŚÉņä£ņØś ĒØĪņłś ĻĖ░ļŖźņØĆ ĒśäņĀĆĒĢśĻ▓ī ļ¢©ņ¢┤ņ¦ĆĻ▓ī ļÉśņ¢┤ ņŗ¼ĒĢ£ ĒØĪņłśļČłļ¤ēņä▒ ņäżņé¼ņÖĆ ĒĢ©Ļ╗ś Ēāłņłśļź╝ ņ£Āļ░£ĒĢśĻ│Ā ĒŖ╣Ē׳ ņ¢┤ļ”░ ļÅ╝ņ¦ĆņŚÉņä£ ļåÆņØĆ ĒÅÉņé¼ņ£©ņØä ļ│┤ņØ┤Ļ▓ī ļÉ£ļŗż.
IHC Ļ▓Ćņé¼ Ļ▓░Ļ│╝ Ļ│ĄņןĻ│╝ ĒÜīņןņŚÉņä£ Ļ┤æļ▓öņ£äĒĢśĻ▓ī ņāüĒö╝ņäĖĒżņŚÉņä£ PEDV ĒĢŁņøÉņØ┤ ļ░£ĒśäļÉśņŚłņ£╝ļ®░ Ļ│Ąņן(82.2%)ļ│┤ļŗżļŖö ĒÜīņן(85.8%)ņŚÉņä£ ļåÆĻ▓ī ļ░£ĒśäļÉśĻ│Ā ņ׳ņŚłļŗż. ļ╣äļĪØ Ļ▓Ćņé¼ĒĢ£ ļÅ╝ņ¦ĆņØś Ļ▓░ņןņŚÉņä£ ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀü ļ│æļ│ĆņØĆ Ļ▒░ņØś Ļ┤Ćņ░░ļÉśņ¦Ć ņĢŖņĢśņ¦Ćļ¦ī, 19.5%ņØś Ļ▓░ņןņŚÉņä£ļÅä ĻĄŁņåīņĀüņØ┤ņ¦Ćļ¦ī PEDV ĒĢŁņøÉņØä Ļ▓ĆņČ£ĒĢĀ ņłś ņ׳ņŚłļŗż. Ļ│╝Ļ▒░ ņŚ░ĻĄ¼ņŚÉ ļö░ļź┤ļ®┤, PEDVņŚÉ ņ×ÉņŚ░Ļ░ÉņŚ╝ļÉśĻ▒░ļéś ņØĖĻ│ĄĻ░ÉņŚ╝ņŗ£Ēé© ļÅ╝ņ¦ĆņŚÉņä£ ņåīņןņØś ņ£Ąļ¬©ņāüĒö╝ņäĖĒżņÖĆ Ļ▓░ņן ņĀÉļ¦ēņŚÉņä£ ĒśĢĻ┤æĒĢŁņ▓┤Ļ▓Ćņé¼ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļ░öņØ┤ļ¤¼ņŖż ĒĢŁņøÉņØä Ļ▓ĆņČ£ĒĢ£ ļ░ö ņ׳ļŗż[2]. ņØ┤ņŚÉ ļ░śĒĢśņŚ¼ Ļ│╝Ļ▒░ ĻĄŁļé┤ ļśÉļŖö ņØ╝ļ│ĖņŚÉņä£ ļ░£ņāØĒĢ£ PEDņØś Ļ▓ĮņÜ░ Ļ▓░ņןņŚÉņä£ļŖö ļ░öņØ┤ļ¤¼ņŖżĻ░Ć Ļ▓ĆņČ£ļÉśņ¦Ć ņĢŖĻ▒░ļéś ņĢäņŻ╝ ņĢĮĒĢśĻ▓ī Ļ▓ĆņČ£ļÉśņŚłļŗż[14,15]. ĻĘĖļ¤¼ļéś ņĄ£ĻĘ╝ 2013ļģä ļ»ĖĻĄŁņŚÉņä£ PEDVļź╝ ņŗżĒŚśĻ░ÉņŚ╝ņŗ£Ēé© ļÅ╝ņ¦ĆņŚÉņä£ļŖö 5ļ¦łļ”¼ ņĀä ļæÉņłśņŚÉņä£ ļ¦╣ņן ļ░Å Ļ▓░ņןņŚÉņä£ ĒĢŁņøÉņØ┤ Ļ▓ĆņČ£ļÉ£ ļ░ö ņ׳ļŗż[6]. ļö░ļØ╝ņä£ ņØ┤ļ▓ł ņĀ£ņŻ╝ļÅäņŚÉņä£ ļ░£ņāØĒĢ£ PEDļŖö Ļ│╝Ļ▒░ ĻĄŁļé┤ņŚÉņä£ ļ░£ņāØĒĢ£ PEDņÖĆ ņן ņĪ░ņ¦ü ļé┤ ļ░öņØ┤ļ¤¼ņŖż ĒĢŁņøÉņØś ļČäĒżĻ░Ć ļŗżļź┤ļ®░, Ļ▓░ņן ļé┤ ĒĢŁņøÉņØś ņĪ┤ņ×¼ļŖö ņØ╝ņĀĢ ņĀĢļÅä ļÅ╝ņ¦ĆņØś ņäżņé¼ļź╝ ļ╣äļĪ»ĒĢ£ ņ×äņāüņ”ØņāüņØä ļŹöņÜ▒ ņŗ¼ĒĢśĻ▓ī ļ¦īļō£ļŖö ņÜöņØĖņ£╝ļĪ£ ņ×æņÜ®ļÉśņŚłņØä Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉ£ļŗż.
PEDņØś ņ×äņāüņ”Øņāü, ļ│æļ”¼ĒĢ┤ļČĆĒĢÖņĀü ļ░Å ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀü ņåīĻ▓¼ņØĆ TGEņÖĆ ļ¦żņÜ░ ņ£Āņé¼ĒĢśĻĖ░ ļĢīļ¼ĖņŚÉ ļæÉ ņ¦łļ│æņŚÉ ļīĆĒĢ£ Ļ░Éļ│äņ¦äļŗ©ņØĆ ļ¦żņÜ░ ņżæņÜöĒĢśļŗż[2,6,13]. ļæÉ ņ¦łļ│æņØä Ļ░Éļ│äĒĢśĻĖ░ ņ£äĒĢ£ ņ¦äļŗ©ĻĖ░ļ▓Ģņ£╝ļĪ£ļŖö ĒśĢĻ┤æĒĢŁņ▓┤Ļ▓Ćņé¼ļ▓Ģ, IHC, ņĀäņ×ÉĒśäļ»ĖĻ▓Į Ļ▓Ćņé¼ļ▓Ģ, ļ░öņØ┤ļ¤¼ņŖżļČäļ”¼ĻĖ░ļ▓Ģ, ĒÜ©ņåīĻ▓░ĒĢ®ļ®┤ņŚŁĒØĪņ░®ņĖĪņĀĢ(enzyme-linked immunosorbent assay) ĻĖ░ļ▓Ģ, RT-PCR ļō▒ņØ┤ ĒÖ£ņÜ®ļÉśĻ│Ā ņ׳ļŗż. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ļÅ╝ņ¦Ć ļČäļ│Ć 153ņĀÉņØä ļīĆņāüņ£╝ļĪ£ RT-PCR Ļ▓Ćņé¼ļź╝ ņłśĒ¢ēĒĢ£ Ļ▓░Ļ│╝, 151ļ¦łļ”¼Ļ░Ć PEDV ņ¢æņä▒ņØ┤ņŚłņ£╝ļéś, ļéśļ©Ėņ¦Ć 2ļ¦łļ”¼ļŖö ņØīņä▒ņØä ļéśĒāĆļé┤ņŚłļŗż. ĻĘĖļ¤¼ļéś ņØ┤ ļæÉ ļ¦łļ”¼ ļÅ╝ņ¦ĆļōżņØĆ ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀüņ£╝ļĪ£ ņåīņן ņ£Ąļ¬©ņØś ņ£äņČĢĻ│╝ ņ£ĄĒĢ® ļ░Å ņāüĒö╝ņäĖĒżņØś ļ│æļ│ĆņØ┤ ņ׳ņŚłņØä ļ┐Éļ¦ī ņĢäļŗłļØ╝, IHC Ļ▓Ćņé¼ Ļ▓░Ļ│╝ PEDV ĒĢŁņøÉ ņ¢æņä▒ņ£╝ļĪ£ ĒīÉļ¬ģļÉśņŚłļŗż. ļö░ļØ╝ņä£ ļÅ╝ņ¦ĆņŚÉ ļ░öņØ┤ļ¤¼ņŖżĻ░Ć Ļ░ÉņŚ╝ļÉ£ Ēøä ņåīņן ņ£Ąļ¬©ņāüĒö╝ņäĖĒżņŚÉ ņåÉņāüņØä ņŻ╝ĻĖ░ļŖö ĒĢśņśĆņ£╝ļéś, ņäĖĒż ļé┤ņŚÉņä£ ņČ®ļČäĒĢ£ ņ¢æņØś ļ░öņØ┤ļ¤¼ņŖż ņ”ØņŗØņØ┤ ņØ┤ļŻ©ņ¢┤ņ¦Ćņ¦Ć ņĢŖņĢä ļČäļ│Ćņ£╝ļĪ£ ļ░öņØ┤ļ¤¼ņŖż ļ░░ņČ£ņØ┤ ņĀüĻ▒░ļéś ņŚåņŚłļŹś Ļ░ÉņŚ╝ ņ┤łĻĖ░ ļŗ©Ļ│äņØś Ļ░£ņ▓┤ļĪ£ ĒĢ┤ņäØļÉĀ ņłś ņ׳ņØä Ļ▓āņ£╝ļĪ£ ĒīÉļŗ©ļÉ£ļŗż. PEDVņØś ņøÉĒśĢņ£╝ļĪ£ ļ░øņĢäļōżņŚ¼ņ¦ĆĻ│Ā ņ׳ļŖö CV777 ļČäļ”¼ņŻ╝ļź╝ Ļ▓ĮĻĄ¼ļĪ£ ņØĖĻ│ĄĻ░ÉņŚ╝ņŗ£Ēé© ļÅ╝ņ¦ĆņŚÉņä£ ĒśĢĻ┤æĒĢŁņ▓┤Ļ▓Ćņé¼ļ▓ĢņØä ņØ┤ņÜ®ĒĢśņŚ¼ ļ░öņØ┤ļ¤¼ņŖż ĒĢŁņøÉņØä Ļ▓Ćņé¼ĒĢ£ Ļ▓░Ļ│╝ Ļ░ÉņŚ╝ Ēøä ņĢĮ 12-18ņŗ£Ļ░äņŚÉ ņåīņן ņāüĒö╝ņäĖĒżņŚÉ Ļ░ÉņŚ╝ņØ┤ ņØ┤ļŻ©ņ¢┤ņ¦ĆĻ│Ā 24-36ņŗ£Ļ░äņŚÉ ņĄ£ļīĆņ╣śņŚÉ ļŗ¼ĒĢśļ®░, ņØ┤Ēøä ļ░öņØ┤ļ¤¼ņŖżņØś ņ”ØņŗØĻ│╝ ĒĢ©Ļ╗ś ņāüĒö╝ņäĖĒżņØś ļ│Ćņä▒Ļ│╝ ņ£Ąļ¬©ņØś ņ£äņČĢņØ┤ ļ░£ņāØĒĢ£ļŗż[2]. ņĄ£ĻĘ╝ ļ»ĖĻĄŁĒśĢ PEDVļź╝ ņØĖĻ│ĄĻ░ÉņŚ╝ņŗ£Ēé© ļÅ╝ņ¦ĆņØś Ļ▓ĮņÜ░ Ļ░ÉņŚ╝ Ēøä 24-48ņŗ£Ļ░ä ĒøäņŚÉ ļČäļ│ĆņØä ĒåĄĒĢ┤ ļ░öņØ┤ļ¤¼ņŖżĻ░Ć ļ░░ņČ£ļÉśņŚłļŗżĻ│Ā ĒĢśņśĆļŗż[6]. RT-PCR Ļ▓Ćņé¼ĻĖ░ļ▓ĢņØĆ PEDVņÖĆ TGEVļź╝ Ļ░Éļ│äņ¦äļŗ©ĒĢśĻĖ░ņŚÉ ļ¦żņÜ░ ņÜ®ņØ┤ĒĢśĻ│Ā ļ»╝Ļ░ÉĒĢ£ ņ¦äļŗ©ĻĖ░ļ▓ĢņØ┤ĻĖ░ļŖö ĒĢśņ¦Ćļ¦ī, ļČäļ│Ć ļé┤ ļ░öņØ┤ļ¤¼ņŖż ļ░░ņČ£ņØ┤ ļ¦żņÜ░ ņĀüņØĆ Ļ░ÉņŚ╝ ņ┤łĻĖ░ ļśÉļŖö ļ¦ÉĻĖ░ņŚÉļŖö Ļ▓ĆņČ£ļÉśņ¦Ć ņĢŖņØä Ļ░£ņŚ░ņä▒ņØ┤ ņ׳ļŗż. ļśÉĒĢ£ IHC ĻĖ░ļ▓ĢņØĆ ļ│æļ”¼ņĪ░ņ¦üĒĢÖņĀü ļ│æļ│ĆņØś Ļ┤Ćņ░░Ļ│╝ ĒĢ©Ļ╗ś ņĪ░ņ¦ü ļé┤ ļ░öņØ┤ļ¤¼ņŖżņØś ņ£Āļ¼┤Ļ╣īņ¦Ć ĒÖĢņØĖĒĢĀ ņłś ņ׳ļŖö ļ¦żņÜ░ ņ£ĀņÜ®ĒĢ£ ņ¦äļŗ©ĻĖ░ļ▓Ģņ£╝ļĪ£ ņ”Øņāü ļ░£Ēśä Ēøä 2ņØ╝ ņØ┤ļé┤ņØś ĻĖēņä▒ĻĖ░ ļÅ╝ņ¦ĆņØś ņן ņĪ░ņ¦ü ņŗ£ļŻīņŚÉ ņĀüņÜ®ĒĢĀ ņłś ņ׳ļŗż. ĒŖ╣Ē׳ ņäżņé¼ ņ”ØņāüņØä ļ│┤ņØ┤ļŖö Ļ░£ņ▓┤ņØś ņŗĀņäĀĒĢ£ ņĪ░ņ¦ü ļśÉļŖö ļČäļ│ĆņØ┤ ņŚåļŖö ņāüĒÖ®ņŚÉņä£ Ēżļź┤ļ¦Éļ”░ Ļ│ĀņĀĢ ņĪ░ņ¦üļ¦īņØ┤ ņ׳ņØä Ļ▓ĮņÜ░ IHC ĻĖ░ļ▓ĢņØĆ PEDV Ļ▓ĆņČ£ņØä ņ£äĒĢ┤ ņ£ĀņÜ®ĒĢśĻ▓ī ņĀüņÜ®ļÉĀ ņłś ņ׳ļŗż. ĻĘĖļ¤¼ļéś IHC ĻĖ░ļ▓ĢļÅä ņé¼Ēøäļ│ĆĒÖöĻ░Ć ņŗ¼ĒĢśĻ▓ī ņ¦äĒ¢ēļÉśņ¢┤ ņ׳Ļ▒░ļéś, ņāüĒö╝ņäĖĒżņØś ĒāłļØĮņØ┤ ĒśäņĀĆĒĢ£ ņåīņןņŚÉļŖö ņ¦äļŗ©ļ▓Ģ ņĀüņÜ®ņŚÉ ņĀ£ĒĢ£ņØ┤ ļö░ļźĖļŗż. ļö░ļØ╝ņä£ ļ░öņØ┤ļ¤¼ņŖżņØś Ļ░ÉņŚ╝ ņŗ£ĻĖ░ņÖĆ Ļ░ÉņŚ╝ļ¤ē, Ļ░ÉņŚ╝ļÉ£ ļÅ╝ņ¦ĆņØś ļ│æļ│Ć ņĀĢļÅä, Ļ▓Ćņé¼ņŚÉ ĒÖ£ņÜ®ĒĢĀ ņłś ņ׳ļŖö ņŗ£ļŻīņØś ņóģļźśņÖĆ ļ│┤Ļ┤Ć ņāüĒā£ ļō▒ņŚÉ ļö░ļØ╝ ņ¦äļŗ©ĻĖ░ļ▓ĢņØś ņĀüņÜ®ņŚÉ ņĀ£ĒĢ£ņØ┤ ļö░ļź╝ ņłś ņ׳ĻĖ░ ļĢīļ¼ĖņŚÉ, RT-PCRĻ│╝ IHC ĻĖ░ļ▓ĢņØä ļ│æņÜ®ĒĢśņŚ¼ ĒÖ£ņÜ®ĒĢśļ®┤ ņóĆ ļŹö ņĀĢĒÖĢĒĢ£ PED ņ¦äļŗ©ņØ┤ ņØ┤ļŻ©ņ¢┤ņ¦ł ņłś ņ׳ņØä Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż.
Ļ│╝Ļ▒░ ĒÆŹĒåĀļ│æņä▒ PEDļĪ£ ņØĖĒĢ£ ņśüĒ¢źņØĆ ĻĘĖļ”¼ Ēü¼ņ¦Ć ņĢŖņĢśņ£╝ļéś, 2010ļģä 10ņøö ņżæĻĄŁ ļé©ļČĆ ņ¦ĆņŚŁ Ēżņ£Ā ņ×ÉļÅłņŚÉņä£ 80-100%ņŚÉ ļŗ¼ĒĢśļŖö ļåÆņØĆ ĒÅÉņé¼ņ£©ņØä ļ│┤ņØ┤ļŖö PEDĻ░Ć ļ░£ņāØĒĢ£ ņØ┤Ēøä ļŗżļźĖ ņĢäņŗ£ņĢä ĻĄŁĻ░ĆļōżļĪ£ ĻĖēņåŹĒ׳ ņĀäĒīīļÉśņŚłņ£╝ļ®░[16], ļ»ĖĻĄŁņŚÉņä£ļŖö 2013ļģä 5ņøö ņĢäņØ┤ņśżņÖĆņŻ╝ņŚÉņä£ ļ│Ė ņ¦łļ│æņØ┤ ņĄ£ņ┤łļĪ£ ļ░£ņāØĒĢ£ Ēøä ņĀäĻĄŁņ£╝ļĪ£ ĒÖĢņé░ļÉśņŚłļŗż[6]. ĻĄŁļé┤ņŚÉņä£ļÅä 2013ļģä ļ¦ÉļČĆĒä░ ņČ®ņ▓Łļé©ļÅäņÖĆ Ļ▓Įņāüļé©ļÅä ņ¦ĆņŚŁņŚÉņä£ PEDĻ░Ć ņ×ćļŗ¼ņĢä ļ░£ņāØĒĢ£ Ēøä ņĀäĻĄŁņĀüņ£╝ļĪ£ ĒÖĢņé░ļÉ£ ņāüĒÖ®ņØ┤ļŗż. ņĀ£ņŻ╝ļÅäļŖö ĻĄŁļé┤ ļé┤ļźÖņ¦ĆņŚŁĻ│╝ ļČäļ”¼ļÉśņ¢┤ ņ׳ļŖö ņä¼ņØ┤ļØ╝ļŖö ĒŖ╣ņłśņä▒ņØä Ļ░Ćņ¦Ćļ®░, 2002ļģä 4ņøöļČĆĒä░ ņĢģņä▒ ļÅ╝ņ¦Ćņ¦łļ│æ ņ▓ŁņĀĢĒÖöļź╝ ļŗ¼ņä▒ĒĢśņŚ¼ ļŗżļźĖ ņ¦ĆņŚŁņ£╝ļĪ£ļČĆĒä░ ļÅ╝ņ¦Ć ņāØņČĢ ļ░Å ļÅ╝ņ¦ĆĻ│ĀĻĖ░ņØś ļ░śņ×ģņØ┤ ņĀäļ®┤ ĻĖłņ¦ĆļÉśņ¢┤ ņ׳Ļ│Ā, ņØ╝ļČĆ ņłśņ×ģņ£Ī ļō▒ņŚÉ ĒĢ£ĒĢ┤ņä£ļ¦ī ļ░śņ×ģņØ┤ ĒŚłņÜ®ļÉśĻ│Ā ņ׳ļŖö ņāüĒā£ņØ┤ļŗż. ļö░ļØ╝ņä£ 2014ļģä 4ņøö ņØ┤Ēøä ļ░£ņāØĒĢśĻ│Ā ņ׳ļŖö ņĀ£ņŻ╝ļÅä ļé┤ PEDņØś Ļ▓ĮņÜ░ Ļ░ĆņČĢ ļō▒ņŚÉ ņØśĒĢ£ ņĀäĒīīļ│┤ļŗżļŖö ĒāĆņŗ£ļÅäņŚÉņä£ ņČ£ņ×ģĒĢśļŖö ņČĢņé░Ļ┤ĆĻ│äņ×É, ņ░©ļ¤ē ļō▒ņŚÉ ņØśĒĢ£ ĻĖ░Ļ│äņĀü ņĀäĒīīņŚÉ ņØśĒĢ┤ ņ£Āņ×ģļÉśņŚłņØä Ļ░ĆļŖźņä▒ņØ┤ ļåÆņØĆ Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉśĻ│Ā ņ׳ļŗż.
Ļ│╝Ļ▒░ ĻĄŁļé┤ņŚÉņä£ ļČäļ”¼ļÉ£ PEDVļŖö CV777 ļČäļ”¼ņŻ╝ņÖĆ ļ¦żņÜ░ ņ£Āņé¼ĒĢ£ Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņÖöļŗż[4]. ĻĘĖļ¤¼ļéś ņĄ£ĻĘ╝ ļåŹļ”╝ņČĢņé░Ļ▓ĆņŚŁļ│ĖļČĆ ņŚ░ĻĄ¼ņŚÉ ļö░ļź┤ļ®┤, ĒĢ£ĻĄŁ ļé┤ļźÖņ¦ĆņŚŁņØś ņ¢æļÅł ļåŹĻ░Ć ļÅ╝ņ¦Ć ņäżņé¼ ļČäļ│ĆņŚÉņä£ ļČäļ”¼ĒĢ£ ļ░öņØ┤ļ¤¼ņŖż 2010ļģä 8Ļ░£, 2012ļģä ļ░Å 2013ļģä 9Ļ░£ ļō▒ ņ┤Ø 17ņŻ╝ņØś PEDVņØś S, open reading frame 3 (ORF3), envelope (E), membrane (M), nucleoprotein (N) ņ£ĀņĀäņ×ÉņŚÉ ļīĆĒĢ£ ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļČäņäØĒĢśņŚ¼ ļČäņ×ÉņāØļ¼╝ĒĢÖņĀü ĒŖ╣ņä▒ņØä ņĪ░ņé¼ĒĢśņśĆļŗż[17]. ĻĘĖ Ļ▓░Ļ│╝ ĻĄŁļé┤ PEDVļŖö Ļ░ü ņ£ĀņĀäņ×Éļ│äļĪ£ Ēü¼Ļ▓ī 2Ļ░£ņØś ĻĘĖļŻ╣(G1 ļ░Å G2)ņ£╝ļĪ£ ļéśļēśņŚłļŗż. PEDV S ņ£ĀņĀäņ×ÉņØś Ļ▓ĮņÜ░ Ēü¼Ļ▓ī ļ░▒ņŗĀņŻ╝Ļ░Ć ņåŹĒĢ£ ĻĘĖļŻ╣(G1)Ļ│╝ ļīĆļČĆļČäņØś ņĢ╝ņÖĖņŻ╝Ļ░Ć ņåŹĒĢśļŖö ĻĘĖļŻ╣(G2)ņ£╝ļĪ£ ļéśļēśĻ│Ā, G2ļŖö ļŗżņŗ£ G2-1Ļ│╝ G2-2ļĪ£ ņäĖļČäļÉśļ®░, ĻĄŁļé┤ļČäļ”¼ņŻ╝Ļ░äņØś ņāüļÅÖņä▒ņØĆ ņĄ£ļīĆ 10% ņØ┤ņāü ņ░©ņØ┤ļź╝ ļéśĒāĆļé┤Ļ│Ā ņ׳ņŚłļŗż. ļśÉĒĢ£ ņŗ£ĒīÉļÉśĻ│Ā ņ׳ļŖö ļ░▒ņŗĀņŻ╝ņÖĆ ĻĄŁļé┤ļČäļ”¼ņŻ╝Ļ░ä Ļ░ü ņ£ĀņĀäņ×Éļ│ä ņāüļÅÖņä▒ ļČäņäØĻ▓░Ļ│╝ ORF3, E, S, N, M ņł£ņ£╝ļĪ£ ņāüļÅÖņä▒ņØ┤ ļé«ņĢśņ£╝ļ®░, S ņ£ĀņĀäņ×ÉņØś Ļ▓ĮņÜ░ 91.8-99.3% (90.0-98.9%)ļĪ£ ņĢĮ 10%ņØś ņ░©ņØ┤ļź╝ ļéśĒāĆļé┤Ļ│Ā ņ׳ņŚłļŗż. ļŹöņÜ▒ņØ┤ ĒĢ┤ņÖĖņŚÉņä£ļÅä 2010ļģä ņØ┤Ēøä ņżæĻĄŁ ļ░Å ļČüļ»Ė ļō▒ņ¦ĆņŚÉņä£ ļ░£ņāØĒĢ£ PEDVļŖö ņä£ļĪ£ ņāüļÅÖņä▒ņØ┤ ļåÆĻ│Ā ļ│æņøÉņä▒ņØ┤ Ļ░ĢĒĢśņŚ¼, 2010ļģä ņØ┤ņĀäņØś PEDVņÖĆ ņ£ĀņĀäņĀü ļ░Å ļ│æņøÉņä▒ņØś ņ░©ņØ┤Ļ░Ć ņ׳ņŚłļŗż[18,19]. ņØ┤ļ¤¼ĒĢ£ PEDVņØś ļ│ĆņØ┤ļĪ£ ņØĖĒĢśņŚ¼ ĻĄŁļé┤ļź╝ ļ╣äļĪ»ĒĢ£ ņÖĖĻĄŁņŚÉņä£ļÅä ĻĖ░ņĪ┤ņŚÉ ņé¼ņÜ®ĒĢśĻ│Ā ņ׳ļŹś ļ░▒ņŗĀņØś ļ░®ņ¢┤ ĒÜ©Ļ│╝ņŚÉ ļīĆĒĢ£ ļ¼ĖņĀ£ņĀÉņØ┤ ņĀ£ĻĖ░ļÉśĻ│Ā ņ׳ļŖö ņŗżņĀĢņØ┤ļŗż. ļö░ļØ╝ņä£ Ļ░üĻĄŁņŚÉņä£ļŖö ņĄ£ĻĘ╝ ļ░£ņāØĒĢśĻ│Ā ņ׳ļŖö PEDņŚÉ ļīĆĒĢ£ ņŗĀņåŹ ņĀĢĒÖĢĒĢ£ ņ¦äļŗ©ĻĖ░ļ▓ĢņØś ĒÖĢļ”Įļ┐Éļ¦ī ņĢäļŗłļØ╝ ņ¦łļ│æ ļ░£ņāØ ĻĖ░ņĀä ļ░Å ņ£ĀĒÜ©ņä▒ņØ┤ ņ׳ļŖö ļ░▒ņŗĀ Ļ░£ļ░£ ļō▒ņØś ņŚ░ĻĄ¼ņŚÉ ļ░Ģņ░©ļź╝ Ļ░ĆĒĢśĻ│Ā ņ׳ļŗż[20,21].
ņØ┤ļ▓ł ņŚ░ĻĄ¼ņŚÉņä£ ļČäļ”¼ļÉ£ ņ┤Ø 12ņŻ╝ņØś PEDV S ņ£ĀņĀäņ×Éļź╝ ļīĆņāüņ£╝ļĪ£ ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļČäņäØĒĢ£ Ļ▓░Ļ│╝ Ļ░ü ļ░öņØ┤ļ¤¼ņŖżņØś ņāüļÅÖņä▒ņØĆ 98.9-99.8%ņŚÉ ļŗ¼ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ĒÖĢņØĖļÉśņŚłļŗż. ļö░ļØ╝ņä£ ņĀ£ņŻ╝ņ¦ĆņŚŁņŚÉņä£ 10ļģä ļ¦īņŚÉ PEDĻ░Ć ņ×¼ļ░£ņāØĒĢśņśĆņ¦Ćļ¦ī, Ļ▒░ņØś ņ£ĀņĀäņĀüņ£╝ļĪ£ ļÅÖņØ╝ĒĢ£ ļ░öņØ┤ļ¤¼ņŖżĻ░Ć ļÅ╝ņ¦ĆņŚÉĻ▓ī ņĀäĒīīļÉśĻ│Ā ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż. ļśÉĒĢ£ PEDVņŚÉ ļīĆĒĢ£ Ļ│äĒåĄĒĢÖņĀü ĒŖĖļ”¼ļź╝ ņ×æņä▒ĒĢ£ Ļ▓░Ļ│╝ ņØ┤ļōż ļ░öņØ┤ļ¤¼ņŖżļŖö ļ¬©ļæÉ 2013-2014ļģä ĒĢ£ĻĄŁ ļé┤ļźÖņŚÉņä£ ļ░£ņāØĒĢ£ PEDV [22] ļ░Å ļČüļ»ĖĒśĢ PEDVņŚÉ ņåŹĒĢśļŖö G2 ĻĘĖļŻ╣ņŚÉ ņåŹĒĢśņśĆļŗż. ĒĢ£ĒÄĖ ĻĄŁļé┤ ļé┤ļźÖņŚÉņä£ ļČäļ”¼ļÉ£ PEDVņØś ņØ╝ļČĆļŖö Ļ░ÖņØĆ G2 ĻĘĖļŻ╣ņŚÉ ņåŹĒĢśņ¦Ćļ¦ī ļ│æņøÉņä▒ņØ┤ ļ╣äĻĄÉņĀü ņĢĮĒĢ£ Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŖö INDELs ĻĘĖļŻ╣ņŚÉ ņåŹĒĢśļŖö Ļ▓āņ£╝ļĪ£ ļ░ØĒśĆņĪīņ£╝ļéś[11], ņØ┤ļ▓ł ņĀ£ņŻ╝ņŚÉņä£ Ļ▓ĆņČ£ļÉ£ 12ņŻ╝ņØś PEDVļŖö INDELs ĻĘĖļŻ╣Ļ│╝ļŖö ļÅÖņ¦łņä▒ņŚÉ ņ░©ņØ┤Ļ░Ć ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ĒÖĢņØĖļÉśņŚłļŗż. ņĄ£ĻĘ╝ Lee ļō▒[23]ņØĆ 2018ļģäņŚÉ ņĀ£ņŻ╝ņŚÉņä£ ļČäļ”¼ļÉ£ PEDVļź╝ G2b PEDVņÖĆ ļ╣äĻĄÉ ļČäņäØĒĢ£ Ļ▓░Ļ│╝ S ņ£ĀņĀäņ×É ļ░Å ņĀäņ▓┤ ņ£ĀņĀäņ▓┤ ņłśņżĆņŚÉņä£ Ļ░üĻ░ü 96.7-98.7% ļ░Å 98.5-99.4% ņāüļÅÖņä▒ņØä ļ│┤ņśĆļŗż. ļö░ļØ╝ņä£ ņ£ĀņĀäņĀü ļ░Å Ļ│äĒåĄ ļ░£ņāØĒĢÖņĀüņ£╝ļĪ£ 2018ļģäņŚÉ ņĀ£ņŻ╝ņŚÉņä£ ļČäļ”¼ļÉ£ PEDVĻ░Ć ĻĖ░ņĪ┤ 2014ļģä ņĀ£ņŻ╝ PEDVņÖĆ ņāüļÅÖņä▒ ļåÆĻĖ░ļŖö ĒĢśņ¦Ćļ¦ī ņŗ£Ļ░äņØ┤ ņ¦ĆļéĀņłśļĪØ ņāüļÅÖņä▒ņØ┤ ļé«ņĢäņ¦ĆĻ│Ā ņ׳ņ¢┤ ļ░öņØ┤ļ¤¼ņŖżĻ░Ć ņ¦äĒÖöĒĢśĻ│Ā ņ׳ņØīņØä ņŗ£ņé¼ĒĢśņśĆļŗż. ļö░ļØ╝ņä£ ņĀ£ņŻ╝ņŚÉņä£ ļ¼ĖņĀ£ņŗ£ļÉśĻ│Ā ņ׳ļŖö PEDVņÖĆ ĻĄŁļé┤ ļé┤ļźÖņ¦ĆņŚŁņŚÉņä£ ļ░£ņāØĒĢśĻ│Ā ņ׳ļŖö PEDVņÖĆņØś ļ│æņøÉņä▒ņØś ņ░©ņØ┤ ļ░Å ļ░öņØ┤ļ¤¼ņŖżņØś ņ£ĀņŚ░ Ļ┤ĆĻ│äļź╝ ļ╣äĻĄÉ ļČäņäØĒĢśļŖö ņŚ░ĻĄ¼ņÖĆ ĒĢ©Ļ╗ś ņĀüņĀłĒĢ£ ļ░▒ņŗĀņŻ╝ņØś ņäĀļ│ä ļ░Å ĒÜ©Ļ│╝ņĀüņØĖ ļ░▒ņŗĀ Ļ░£ļ░£ņØä ĒåĄĒĢśņŚ¼ PEDļĪ£ ņØĖĒĢ£ ņ¢æļÅł ļåŹĻ░ĆņØś Ēö╝ĒĢ┤ļź╝ ņĄ£ņåīĒÖöĒĢśļŖöļŹ░ ļģĖļĀźņØä Ļ▓ĮņŻ╝ĒĢ┤ņĢ╝ ĒĢĀ Ļ▓āņ£╝ļĪ£ ĒīÉļŗ©ļÉ£ļŗż.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print